Informationsbehandling er vigtig inden for alle videnskabelige områder. I molekylærbiologi er den centrale dogme, der først blev opfundet af Francis Crick (Crick, 1958, 1970), en klassisk rygrad i levende celler til fundamentalt at udføre processer fra celledeling til død gennem DNA-, RNA- og proteininformationsveje. Mere specifikt beskriver det centrale dogme overførslen af sekvensinformation under DNA-replikation, transkription til RNA og translation til aminosyrekæder, der danner proteiner. Samtidig siger det også, at information ikke kan strømme fra protein til protein eller nukleinsyre.
Siden fremkomsten af systemiske og høje kapacitetsmetoder i løbet af de sidste to årtier, er disse brede trin, som ikke inkluderer komplekse lovgivningsmæssige detaljer, er kommet under intens kontrol. De manglende regulatoriske funktioner, såsom DNA-korrekturlæsning / reparationsmekanismer og alternativ splejsning af præ-mRNA, introducerer flere mellemliggende trin. Disse yderligere trin forstyrrer dogmens nøgletrin og ændrer sandsynligvis informationsdynamikken. Derudover synes epigenetik eller den rolle, som kromatinstrukturer, DNA-methylering og histonmodifikationer spiller, også at gå imod dogmens enkle veje (Shapiro, 2009; Luco et al., 2011). Proteinsplejsning, eller et proteins (inteins) evne til at ændre sin egen sekvens, opdaget i nyere tid (Volkmann og Mootz, 2012) og prioner, som ændrer andre proteinsekvenser (Prusiner, 1998), omgår informationsoverførselsvejen for dogme. Andre undersøgelser rapporterede om fejl eller uoverensstemmelser mellem RNA-sekvenser og deres kodende DNA (Hayden, 2011; Li et al., 2011). Samlet set rejser disse data tvivl om gyldigheden af det centrale dogme i forbindelse med nutidens videnskab og sætter derfor spørgsmålstegn ved enkelheden af lineær informationsstrøm (DNA til RNA og RNA til protein).
For at sætte tingene i perspektiv kræver vi analytiske værktøjer, der undersøger bekymringerne eller uoverensstemmelser vedrørende den mangeårige teori. En enkel, men alligevel meget nyttig teknik til søgning i globale egenskaber i datasæt med høj gennemstrømning er statistisk korrelationsanalyse, som er blevet bredt og med succes brugt til at observere mønstre i komplekse systemer som vejret (Stewart, 1990), aktiemarkeder (Lo og MacKinlay , 1988) og kosmologi (Amati et al., 2008). Der er flere slags korrelationsanalyser, der evaluerer både lineære (fx Pearson-produktmoment) og ikke-lineære (f.eks. Spearmans rang, gensidig information) afhængigheder (Steuer et al., 2002; Rosner, 2011). , er Pearson-produkt-øjeblik-korrelationsanalysen blevet den mest populære på grund af dens evne til at vise organisationsstruktur i den enkleste form.
I biologi har der været adskillige værker, der har studeret sammenhængene i mRNA og proteinekspressionsdata (se nedenfor og tabel 1). I teorien, når to prøver, der indeholder højdimensionale (såsom mikroarray og proteomiske) data, sammenlignes, giver korrelationsanalyserne et mål for afvigelse fra enhed som en kilde til forskel mellem prøverne Kort sagt viser to prøver med identisk og fuldstændig ikke-identisk information henholdsvis enhed (R2 = 1) og nul (R2 = 0) korrelation.
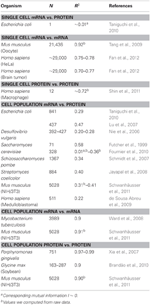
Tabel 1. mRNA og protein expr essionskorrelationer i forskellige organismer.
Perfekt korrelation (R2 = 1) er en idealiseret situation, der er langt fra virkeligheden, som teknisk eller eksperimentel støj alene interfererer og reducerer korrelation. Desuden har de seneste år fremhævet eksistensen af biologisk støj: undersøgelserne af individuelle celler og molekyler har vist stokasticitet i genekspressionsdynamik på grund af den kombinatoriske effekt af lave molekylære kopital og den kvantale karakter af promotordynamik (Raj og van Oudenaarden, 2009; Eldar og Elowitz, 2010). På den anden side viser klonale populationer af celler heterogenitet i niveauerne af et givet proteinekspression pr. Celle til enhver målt tid (Chang et al., 2008). Tilsammen er stokasticitet og heterogenitet afgørende for at producere celle skæbnespredning, fænotypiske variationer og forstærkning af intracellulære signaler (Locke et al., 2011; Selvarajoo, 2012).
De stokastiske udsving eller iboende støj forårsager ekspressionen af en molekylær art til at variere i tid og mellem celler, hvilket fører til ukorrelerede reaktioner (Elowitz et al., 2002). Dette er især fremtrædende for mRNAer og proteiner med lave antal kopier. Således kan korrelationen mellem prøver (celler) sænkes på grund af iboende støj (figur 1A). Andre kilder til biologisk støj på grund af ekstrinsiske faktorer inkluderer variation i cellestørrelse, molekylære kopiantal og miljøudsving mellem individuelle celler.Disse faktorer fordrejer det deterministiske centrale dogme og ændrer sandsynligvis stærke korrelationer til svagere (figur 1B).
En nylig undersøgelse sammenlignede Escherichia coli mRNA og proteinudtryk mellem individuelle celler på et enkelt molekyleniveau og tilvejebragte et scenario, der dybt sætter spørgsmålstegn ved det centrale dogme. Taniguchi et al. (2010) afslørede, at der ikke er nogen sammenhæng (R2 ~ 0) mellem individuelt tufA mRNA og proteinniveauer i enkeltceller. Navnlig konkluderede de, at manglen på korrelation sandsynligvis skyldes forskelle i mRNA- og proteinlevetid. Selv om dette er en plausibel forklaring, Taniguchi et al. var omhyggelig med ikke at afkræfte den langvarige hypotese ved at hævde, at tidsgennemsnit for mRNA-niveauer skulle korrelere med proteinniveauer. Der var imidlertid ingen beviser, der viste, at dette er det egentlige tilfælde, og når vi vurderede ikke-lineære afhængigheder ved hjælp af gensidig information (Steuer et al., 2002; Tsuchiya et al., 2010) i Taniguchi et al. datasæt, fandt vi, at resultatet var ikke-afhængigt, dvs. I ~ 0. Dette bekræfter, at mRNA til proteinekspressioner mellem individuelle celler på enkelt molekyleniveau er klart ikke-relaterede. Når man zoomer på enkeltmolekyle niveau i korrelationsplottet, er det tydeligt, at deres parvise korrelationer er svage (figur 1A, indsæt, til illustration).
Især på cellepopulationniveau, Taniguchi et al. var i stand til at vise relativt høj korrelation mellem mRNA og proteinekspressioner med R2 = 0,29 (figur 2A). Faktisk en anden uafhængig undersøgelse af Lu et al. (2007) for E. coli-populationen viste også relativt høj korrelation (R2 = 0,47). Lignende analyser udført på Saccharomyces cerevisiae (Futcher et al., 1999), murin NIH / 3T3 fibroblast (Schwanhäusser et al., 2011) og flere andre cellepopulationer (Nie et al., 2006; Schmidt et al., 2007; Jayapal et. al., 2008; de Sousa Abreu et al., 2009) viste alle sammenhængende strukturer mellem transkriptomdækkende og proteomdækkende udtryk (tabel 1). Så hvorfor er der ingen sammenhæng mellem individuel mRNA og proteinudtryk i enkeltceller, mens der på populationsniveau observeres kollektive forhold mellem storskala mRNA og proteinudtryk?
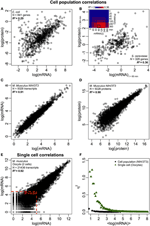
Figur 2. Omics-wide expression correlations. Cellepopulationer: mRNA-proteinkorrelationer i (A) E. coli (Taniguchi et al., 2010) og (B) S. cerevisiae (Fournier et al., 2010) mellem mRNA-udtryk ved t = 60 min og proteinekspressioner ved t = 360 min. Indsæt: korrelationsmatrix mellem alle tidspunkter viser en forsinket stigning i korrelationer mellem mRNA og proteiner. (C) mRNA og (D) proteinudtryk mellem to prøver af murine NIH / 3T3-celler (Schwanhäusser et al., 2011). Enkeltceller: (E) mRNA-udtryk mellem to oocytter (Tang et al., 2009). De røde stiplede linjer angiver regionerne med lave mRNA-udtryk (log (mRNA) < 5). (F) Støj (η2) versus log (mRNA-udtryk) for cellepopulation (NIH / 3T3, sorte prikker, Schwanhäusser et al., 2011) og enkeltceller (oocytter, grønne trekanter, Tang et al., 2009). Hver prik repræsenterer værdien for en gruppe af P = 100 mRNAer. η2 er næsten nul for cellepopulationen for alle mRNA-udtryk. For enkeltceller er η2 højest for mRNAer med de laveste kopital og nærmer sig nul for højere antal kopier.
Vi mener der er to hovedårsager til forskellene. For det første, som tidligere nævnt, reducerer støj, hvad enten den er biologisk eller ikke-biologisk, korrelationen. Da analyser på enkeltceller har vist betydningen af stokasticitet og variation, er disse effekter afgørende for at reducere enkeltcellekorrelationer. På ensemble niveau, når celler samples i en population, reduceres den samlede (iboende + ydre) støj, da tilfældig støj annulleres over alle spektrum af molekylære udtryk (figur 1C-F) for at afsløre gennemsnitlig respons og selvorganisering ( Karsenti, 2008; Selvarajoo, 2011; Hekstra og Leibler, 2012; Selvarajoo og Giuliani, 2012). Derfor opstår en god grad af mRNA-proteinekspressionskorrelation. For det andet blev individuel mRNA-proteinekspressionskorrelation sammenlignet på tværs af adskillige celler for enkeltcelleundersøgelsen (Taniguchi et al., 2010). I cellepopulationsundersøgelser foretages sammenligningen imidlertid i sin helhed på tværs af tusinder af mRNAer og proteiner over flere størrelsesordener større end det ekspressionsområde, der findes for et enkelt molekyle mellem celler. Dette fører derfor til højere korrelationer på populationsniveau, da effekten af enkeltmolekylære variationer bliver ubetydelig.
På trods af at der observeres korrelerede strukturer for cellepopulationer, er der konkrete årsager til den store afvigelse fra perfekt korrelation.Som tidligere nævnt er et nøglepunkt, at mRNAer og proteiner sekventielt er placeret med flere manglende processer, ikke repræsenteret i det centrale dogme. Tilføjelse af de manglende mellemprodukter langs en biokemisk vej vil medføre en mærkbar forsinkelse i informationsstrømmen (Selvarajoo, 2006, 2011; Piras et al., 2011), og korrelationen mellem dem kan lide som et resultat. Dette kunne også være en del af det faktum, som Taniguchi et al. at mRNA og proteinekspressioner har forskellige levetider. Navnlig understøttes denne postulation i et nyligt arbejde med S. cerevisiae behandlet med Rapamycin, der viste de tidsmæssige korrelationer af mRNA-proteinekspression var oprindeligt lave, R2 = 0,01 ved 40 minutter, ikke desto mindre over 360 minutter efter forstyrrelse steg korrelationen, R2 = 0,36 (Fournier et al., 2010, figur 2B). Dataene indikerer, at ved kemisk forstyrrelse afviger det oprindelige respons mellem mRNA og proteinudtryk på grund af tidsforsinkelse og forskellige kinetiske mekanismer mellem dem såvel som sekundære effekter såsom autokrin eller parakrin signalinterferens (Shvartsman et al., 2002; Isalan et al., 2008). Når virkningerne af forstyrrelsen dæmpes over tid, opstod genopretningen af korrelationer.
For yderligere at kontrollere postulationen, at sekventielle forsinkelsesprocesser eller forskellige levetider er afgørende for at mindske mRNA-protein-korrelationer, sammenlignede vi R2 mellem samme molekylære art af det centrale dogme (f.eks. mellem mRNA og mRNA) i cellepopulationer og enkeltceller. Den transkriptomdækkende mRNA-mRNA-ekspressionskorrelation mellem replikater af NIH / 3T3 (Schwanhäusser et al., 2011) (figur 2C) og Mycobacterium tuberculosis (Ward et al., 2008) cellepopulationsprøver er begge meget høje med R2 > 0.9 (tabel 1). Sådanne stærke korrelationer observeres også mellem populationsprøver for protein-protein-udtryk i NIH / 3T3-celler (Schwanhäusser et al., 2011) (figur 2D), Porphyromonas gingivalis (Xia et al., 2007) og Glycine max (Brandão et al. , 2010) (tabel 1). Da disse data, der sammenligner de samme arter, giver meget høje korrelationer, kan det tænkes, at de sekventielle forsinkelsesprocesser eller forskellige levetider er ansvarlige for at sænke populationsniveauets korrelationsstrukturer mellem mRNA og proteinudtryk.
I enkelt murine oocytter ( Tang et al., 2009), når man sammenligner hele mRNA – mRNA-udtryk, observeres en stærkt korreleret struktur (R2 = 0,92, figur 2E). Fokuserer imidlertid kun på lavt udtrykte mRNAer (med logaritmiske udtryk < 5), sænker den stokastiske støj den parvise korrelation ganske dramatisk (R2 < 0,54). For at undersøge dette resultat vurderede vi støj, η2 = σ2XY / μ2XY, på tværs af hele mRNA-udtryk (figur 2F). Vi bemærkede, at η2 er højest for de laveste udtryk på grund af den udtalt effekt af stokastiske udsving i sammenligning med deres udtryk og nærmer sig nul for højere udtryk, hvor sådan støj bliver mindre signifikant (Piras et al., 2012). For cellepopulation observeres som forventet næsten nul støj over hele ekspressionsområdet på grund af annullering af tilfældig støj (figur 1E, F).
Meget korrelerede strukturer for hele mRNA – mRNA-udtryk var også rapporteret for enkelt kræftcelle (Fan et al., 2012), omend mindre signifikant med R2 ~ 0,7 (tabel 1). Yderligere viste sammenligning af protein-protein-udtryk i LPS-stimulerede humane makrofager også høje korrelationer, R2 ~ 0,72 (Shin et al., 2011) (tabel 1). Selvom der ikke er nogen sammenhæng mellem individuelle mRNA-proteinekspressioner i enkeltceller, er den store eller omiske-store korrelation mellem samme molekylære arter i enkeltceller meget høj.
Således uanset om det er enkeltceller eller cellepopulationer , indikerer de omiske brede data, at korrelationerne mellem de samme molekylære arter (mRNA vs. mRNA og protein vs. protein) er mærkbart højere end mellem forskellige arter (mRNA vs. protein). Dette afspejler det faktum, at selvom tidsforsinkelsesprocesser og forskellige levetider er nøglen til at reducere korrelationer, er disse mekanismer ikke tilstrækkelige til at understøtte den manglende korrelationsstruktur, der er observeret mellem individuelle cellers individuelle transkription til proteinudtryk.
Så langt har vi ved at undersøge store udtryk for mRNAer og proteiner fra forskellige cellulære systemer vist, at korrelationsstrukturer opstår i en global skala. Imidlertid afslører korrelationsanalyserne kun forbindelsen mellem to testede prøver og viser ikke retningen af For at det centrale dogme skal være gyldigt på global skala, skal den samlede informationsstrøm være fra DNA til proteiner. Sådan strøm af information er blevet demonstreret af utallige andre undersøgelser, der involverer forstyrrelse af cellerne i cellerne og overvågning af den resulterende dynamik af transkriptionsfaktorer, der binder til DNA og induktion af genekspressioner i stor skala (figur 3A).For eksempel i tilfælde af LPS-stimulerede immunceller er det blevet påvist, at aktiveringen af transkriptionsfaktoren NF-KB finder sted omkring 15 minutter (Liu et al., 1999), induktion af dens nedstrømsgener ved ca. min (Liu et al., 1999; Xaus et al., 2000; Selvarajoo et al., 2008) og translationen af de tilsvarende proteiner i området 60-90 min (Kawai et al., 1999; Xaus et al. ., 2000) (figur 3B). En sådan sekventiel retning af den samlede transkription til informationsflow for translation observeres også for bakteriesystemer, såsom E. coli, på cellepopulationsniveau (Golding et al., 2005).

Figur 3. Informationsstrømmen af central dogme. (A) Skematisk af LPS / TLR4-induceret TNF-ekspression via transkriptionsfaktor NF-KB og tnf-gen efter lineær informationsstrøm. (B) Eksperimentelle tidsmæssige profiler af promotorbindende aktivitet af NF-KB (øvre paneler), tnf (midterste paneler) og TNF (nedre paneler) udtryk på cellepopulation. (C) Skematiske temporale profiler af promotordynamik, mRNA og proteinudtryk på enkeltcelleniveau (Raj og van Oudenaarden, 2009).
Alternativt afslører undersøgelser ved enkeltcelleopløsning tilfældige udsving over den lineære informationsstrøm: transkriptionsfaktorerne, der er bindende til DNA-promotorregioner, er quantal, hvilket resulterer i sprængningsadfærd for mRNA-transkriptionen og inducerer derefter variation i proteinoversættelsen selv mellem identiske celler (figur 3C) (Raj og van Oudenaarden, 2009; Eldar og Elowitz, 2010; Locke et al., 2011; Hekstra og Leibler, 2012; Selvarajoo, 2012). Som et resultat på ethvert bestemt tidspunkt er det individuelle molekylære respons for enkeltceller temmelig støjende sammenlignet med befolkningens gennemsnitlige skala (Selvarajoo, 2011).
Konklusioner
Eksemplerne vist i dette papir fremhæver forskellene i rækkefølgen af korrelationsværdier observeret mellem arter i det centrale dogme i forhold til cellepopulationer og enkeltceller. De statistiske analyser fra cellepopulationer tegner et billede af, at ekspressionskorrelationen mellem de samme molekylarter er meget høj og mellem arter er moderat høj. Skønt enkeltcellekorrelationer mellem de samme arter er sammenlignelige med cellepopulationer, viste de en bredere spredning i deres udtryksdiagrammer på grund af den udtalt effekt af biologisk støj, især for udskrifter med lave kopital. Især bliver de enkelte celler “parvis korrelation nul for individuelle molekyler (Taniguchi et al., 2010). Faktisk er stokastiske udsving og variabilitet i molekylære udtryk kendt for at være funktionelle til at generere beslutning om celleskæbne og vælte cellulære tilstande (Losick og Desplan, 2008; Eldar og Elowitz, 2010; Kuwahara og Schwartz, 2012). Vi mener, at de stærke omics-brede korrelationer forekommer som et resultat af stramme gen- og proteinregulerende netværk på tværs af tusinder af molekyler (Barabási og Oltvai, 2004; Karsenti , 2008), hvilket resulterer i fremvoksende gennemsnitlige responser.Analyser af lille antal eller individuelle molekyler kan ikke korrelationsstrukturen observeres.
Samlet set kan det tænkes, at visning af informationsstrømmen af enkelt DNA til protein vil sætte spørgsmålstegn ved det centrale dogme da responset fra hvert molekyle på et hvilket som helst tidspunkt sandsynligvis ikke vil korrelere. Imidlertid antyder observationen af gennemsnitlig deterministisk respons globalt, at nettets ligevægt i genet IC-oplysninger forbliver yderst til højre for stierne. Derfor bør det centrale dogme ses som en makroskopisk cellulær informationsstrøm i en omisk bred skala og ikke på et enkelt gen til protein-niveau. Som sådan mener vi, at dets enkelhed fortsat vil forblive som en af de mest indflydelsesrige teoretiske søjler i levende systemer.
Erklæring om interessekonflikt
Forfatterne erklærer, at forskningen blev udført i fraværet af kommercielle eller økonomiske forhold, der kan fortolkes som en potentiel interessekonflikt.
Anerkendelser
Kentaro Hayashi takkes for kommentarer. Forskningsfonden i Tsuruoka city og Yamagata Prefecture værdsættes for deres støtte.
Crick, F. (1958). Om proteinsyntese. Symp. Soc. Exp. Biol. 12, 139–163.
Pubmed Abstract | Pubmed fuld tekst
Crick, F. (1970). Molekylærbiologisk central dogme. Nature 227, 561-563.
Pubmed Abstract | Udgivet fuld tekst
Hayden, E. C. (2011). Bevis for ændret RNA vækker debat. Nature 473, 432.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Hekstra, D. R. og Leibler, S. (2012). Beredskab og statistiske love i replikerede mikrobielle lukkede økosystemer. Cell 149, 1164–1173.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Kuwahara, H. og Schwartz, R. (2012).Stokastisk steady state gevinst i en genekspressionsproces med mRNA-nedbrydningskontrol. J. R. Soc. Interface 9, 1589-1598.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Nie, L., Wu, G. og Zhang, W. (2006). Korrelation af mRNA-ekspression og protein overflod påvirket af flere sekvensfunktioner relateret til translationel effektivitet i Desulfovibrio vulgaris: en kvantitativ analyse. Genetik 174, 2229-2243.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Prusiner, S. B. (1998). Prions. Proc. Natl. Acad. Sci. U.S.A. 95, 13363–13383.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Rosner, B. (2011). Grundlæggende om biostatistik. 7. udgave. Boston, MA: Duxbury Press.
Selvarajoo, K. (2006). Opdage differentieringsaktivering af de vejafgiftslignende receptor 4 signalveje i MyD88 knockouts. FEBS Lett. 580, 1457–1464.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Selvarajoo, K. (2011). Makroskopisk lov om bevarelse afsløret i populationsdynamikken ved vejlignende receptorsignalering. Cell Commun. Signal. 9, 9.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Selvarajoo, K. (2012). Forståelse af multimodale biologiske beslutninger fra enkeltcelle og populationsdynamik. Wiley Interdiscip. Præst Syst. Biol. Med. 4, 385-399.
Pubmed Abstract | Udgivet fuld tekst | CrossRef fuldtekst
Stewart, T. R. (1990). En nedbrydning af korrelationskoefficienten og dens anvendelse til analyse af prognosefærdigheder. Vejrudsigt. 5, 661-666.