Introduktion
Gaben er en ufrivillig sekvens af mundåbning, dyb inspiration, kort apnø og mere eller mindre langsom udløb (Baenninger, 1997; Walusinski og Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Når man fremkalder, kan et gaben ikke undertrykkes fuldstændigt. Derfor er det blevet defineret som et stereotype eller reflekslignende mønster (Lehmann, 1979; Provine, 1986). I Homo sapiens er der fremsat adskillige hypoteser med variabel understøttelse for at forklare mekanismer og funktioner til spontan gaben, såsom iltning (åndedrætsfunktion forårsaget af hypoxi), stressrelateret adfærd (forårsaget af ophidselse) eller termoregulering (forårsaget af hypertermi Guggisberg et al., 2010; Massen et al., 2014; Gallup og Gallup, 2019). At være et fysiologisk respons, kan gaben påvirkes af interne og eksterne faktorer som tidspunktet på dagen (Giganti og Zilli, 2011) eller intrakraniel / hjernetemperatur (Gallup og Eldakar, 2013).
Gabe kan være selvstyret og / eller vises for andre (Moyaho et al., 2017; Palagi et al., 2019). I humane og ikke-humane primater, afhængigt af arten, når gabning vises for andre, kan den kommunikere trussel (Troisi et al., 1990; Deputte, 1994) og / eller fysiologiske og adfærdsmæssige ændringer (Provine et al., 1987 ; Leone et al., 2015; Zannella et al., 2015). Hos mennesker er gaben et socialt moduleret respons, fordi det kan hæmmes af faktisk – og ikke virtuel – social tilstedeværelse (Gallup et al., 2019), og fordi en gab kan udløses af andres gab som et resultat af et kendt fænomen. som smitsom gaben (Provine, 1989, 2005). Gapsmitte kan fremkaldes, selvom gaben høres, men ikke ses (Arnott et al., 2009; Massen et al., 2015).
Hos mennesker er deres fylogenetisk nærmeste abeart (chimpanser: Pan paniscus) ; bonobos: Pan troglodytes) og den afrikanske abe Theropithecus gelada, smitsom gaben er ikke kun til stede (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; men se: Amici et al., 2014) men også socialt moduleret, fordi gabresponsen er højest, når visse kategorier af individer er involveret (f.eks. pårørende, gruppemedlemmer, dominanter; Palagi et al., 2009; Campbell og de Waal, 2011, 2014; Norscia og Palagi, 2011; Demuru og Palagi, 2012; Massen et al., 2012). To hovedargumenter er blevet præsenteret for at forklare denne sociale asymmetri i smitsom gaben, som er grupperet i to hovedhypoteser: Emotional Bias Hypothesis (EBH), der forbinder smitsom gaben med følelsesmæssig overførsel, og Attentional Bias Hypothesis (ABH), der betragter smitsom gaben som en motorisk reaktion, der er genstand for forskelle i top-down opmærksomhedsprocesser (Palagi et al., 2020).
EBH forudsiger, at den sociale asymmetri, der observeres i gabe-smittehastigheder, afspejler forskelle i de forskellige social binding, en proxy for følelsesmæssig binding, mellem individer. Denne hypotese understøttes af beviser for, at smittehastigheder i gabe følger en empatisk tendens (sensu Preston og de Waal, 2002), idet den er højest mellem individer, der deler en stærkeste følelsesmæssig bånd. Specifikt fandt Norscia og Palagi (2011), at smittefrekvenser hos mennesker er størst som svar på familie og venner end som svar på bekendte og fremmede. Hos voksne chimpanser er gaben smitte højere mellem i gruppen sammenlignet med medlemmer ude af gruppen (Campbell og de Waal, 2011), og i bonobos er gaben smittehastigheder størst mellem individer, der forbinder mere med hinanden (Demuru og Palagi, 2012). I en sammenlignende undersøgelse, der inkluderer både mennesker og bonoboer, har Palagi et al. (2014) fandt ud af, at gabenes smittehastigheder blev påvirket af forholdskvaliteten mellem individer mere end af arten, som forsøgspersonerne tilhørte. Derudover øges smitte hos mennesker med alderen, når evnen til at identificere andres følelser stiger og falder med alderen, når en sådan evne aftager (Wiggers og van Lieshout, 1985; Anderson og Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen og Anderson, 2011; Bartholomew og Cirulli, 2014). Gapsmittehastigheder øges fra barndom til voksen alder også hos chimpanser (Madsen og Persson, 2013).
ABH forudsiger, at den sociale asymmetri, der observeres i gabesmitte, kan skyldes forskelle i social, visuel opmærksomhed (Massen og Gallup, 2017). Især ville de højeste niveauer af smitsom gab skyldes den ekstra top-down, selektive visuelle opmærksomhed, der er rettet mod individer, der er mere relevante for observatøren, såsom velkendte emner, da det forekommer hos mennesker og gelader eller dominanter, da det forekommer i chimpanser eller bonoboer (Yoon og Tennie, 2010; Massen et al., 2012; Massen og Gallup, 2017).Ifølge Massen og Gallup (2017) ville ABH blive bakket op af den eksisterende evidens for den forskellige visuelle detektion og visuelle opfattelsesfulde kodning af ansigter hos velkendte og / eller gruppeemne i forhold til ukendte (fx Buttle og Raymond, 2003 ; Ganel og Goshen-Gottstein, 2004; Jackson og Raymond, 2006; Michel et al., 2006).
I denne undersøgelse analyserede vi data om gabning, der blev indsamlet over 9 år på mennesker i deres naturlige omgivelser og vi ekstrapolerede de tilfælde, hvor det gab, der udsendes af et emne, kunne høres, men ikke ses af en potentiel responder (auditiv gab). Ved kun at overveje de tilfælde, hvor den visuelle indikation af den gabende stimulus ikke kunne detekteres, verificerede vi, om den sociale asymmetri, der tidligere blev observeret i gabenesmittehastigheder, varede eller ej. Navnlig testede vi følgende alternative forudsigelser afledt af de to hypoteser, der blev præsenteret ovenfor (EBH og ABH).
Forudsigelse 1a: ifølge EBH er hastighederne for gabesmitte påvirket af styrken af inter -individuel social bånd – en proxy for den følelsesmæssige bånd – i sig selv og ikke ved en anden top-down, selektiv visuel opmærksomhed på især bestemte individer. Hvis denne hypotese understøttes, forventer vi at observere den sociale skævhed også, når den visuelle indikation af den gabende stimulus er udelukket, og satserne for auditiv smitsom gabning vil være højere mellem stærkt bundet sammenlignet med svagt bundet individer.
Forudsigelse 1b: ifølge ABH ville de højere niveauer af gaben smitte mellem stærkt bundet sammenlignet med svagt bundet individer være knyttet til den nærmeste top-down, selektive visuelle opmærksomhed, som individer lægger på individer, der er relevante for dem, fx familie og venner. Hvis denne hypotese understøttes, skal den sociale bias, der observeres i gaben-smitten, forsvinde, når kun auditive gab overvejes, fordi den visuelle signal ikke kan overværes af den potentielle responder.
Materialer og metoder
Dataindsamling og operationelle definitioner
Til denne undersøgelse betragtede vi de vokaliserede gab udsendt af et emne, der kun kunne høres – men ikke ses – af en potentiel responder (herefter: auditiv gab). Emitteren og de potentielle respondenter måtte være i et område på ≤5 m. Vokaliserede gab involverede brugen af stemmefold og de gabber, der kun involverede tung inspiration / udløb, blev ikke betragtet som vokaliserede; 294 tilfælde af hørselsgab blev ekstrapoleret fra et datasæt på i alt 2001 gabeanfald indsamlet i løbet af 9 år – fra 2010 til 2019 – ved hjælp af prøvetagningsmetoden for alle forekomster (Altmann, 1974). Specifikt blev auditive gabe indsamlet fra november 2010 til maj 2019 fra 05.30 til 02.30 om humane kaukasiske forsøgspersoner i alderen 18 til 77 under deres rutinemæssige aktiviteter, fx på arbejdspladser, over måltider, under sociale møder osv. ., med emnerne uvidende om at blive observeret og i mangel af nogen tydelig ekstern kilde til angst. Den auditive gabedatabase omfattede 193 gyperpotentiale responder dyader. Afhængig af situationen blev oplysningerne registreret, ubemærket gennem alfanumeriske koder og indtastet direkte i beregningsark, skrevet i mobiltelefoner eller skrevet på papir og derefter indtastet i beregningsark til efterfølgende udarbejdelse. Grundlæggende oplysninger som alder og forholdet mellem mennesker var kendt af forfatterne. De potentielle respondenter blev kodet som i den ikke-synlige tilstand, når deres hoved blev drejet 180 ° i forhold til aftrækkeren, eller når en fysisk, synsblokerende forhindring var til stede, der forhindrede den potentielle responder i at se udløserens ansigt og krop. Trigger og responder blev aldrig helt isoleret (fx i to separate rum med lukkede døre) fra hinanden. Den sociale nærhed blev samlet på fire niveauer: 0 = fremmede, der aldrig havde mødt før; 1 = bekendte, der udelukkende delte et indirekte forhold baseret på et tredje eksternt element, det vil sige arbejdspligt (kolleger) eller venner til fælles (venner af venner); 2 = venner, ikke-relaterede personer, der deler et direkte forhold, der ikke udelukkende er relateret til et tredje eksternt element; 3 = faste partnere og pårørende (r ≥ 0,25). Tidligere litteratur rapporterer, at gabesvar kan fremkaldes inden for 5 minutter efter at have set en andens gabe (triggerens gab) (Provine, 1986) med et maksimum i det første minut (Provine, 2005; Palagi et al., 2014). Litteratur rapporterer også, at der fra det fjerde minut er der den højeste sandsynlighed for autokorrelation (hvilket betyder, at tilstedeværelsen af et gab udført af et motiv ved t0 øger sandsynligheden for at have en anden gab af det samme emne ved t (0 + X), hvor X er den stigende tidsenhed; Kapitány og Nielsen, 2017). Derfor betragtede vi de gabesvar, der optræder inden for et tidsvindue på 3 minutter fra det gab, der udsendes af udløseren.For yderligere at reducere autokorrelationsforstyrrelsen registrerede vi i tilfælde af en kæde af gaber, der udsendes af udløseren (flere gaber udsendt i 3-minutters tidsvinduet) som svar kun den første gabe, der blev udført efter opfattelsen af den sidste gab. Vi kodede en gab som “spontan”, når intet andet emne havde gaben i de 5 minutter forud for gabenhændelsen.
Statistiske analyser
Til analyserne blev følgende variabler overvejet: forekomst af smitte, kodet som: 1 = tilstedeværelse, 0 = fravær; den sociale bånd blev indtastet med de fire niveauer defineret ovenfor (0 = fremmede; 1 = bekendte; 2 = venner; 3 = familie); trigger og observatørs køn blev mærket som : M = mand, F = kvinde; udløserens og responderens aldersklasser blev kodet som følger: yo = ungdom (18-24 år); annonce = voksen (25-64 år gammel); se = senior (ovenfor 65 år gammel) (Statistics Canada, 2009); tidslukerne blev kodet som følger: 1 = 05:30 – 09:00; 2 = 09:01 – 12:30; 3 = 12: 31–16: 00 pm; 4 = 16: 01-19: 30 pm; 5 = 19: 31-23: 00 pm; 6 = 23: 01–02: 30 (Giganti og Zilli, 2011). Databasen (se Supplerende datablad) inkluderede 84 mænd, 69 kvinder, 16 unge (åå), 122 voksne (annonce) og 15 ældre (se). For at teste om forekomsten af gabe smitte blev påvirket af faktorernes bånd (0 = fremmede; 1 = bekendte; 2 = venner; 3 = pårørende), udløserens køn (Trigger_sex), responderens køn (Responder_sex), triggerens aldersklasse (Trigger_ageclass) og responderens aldersklasse (Responder_ageclass) og tidsrum (fra 1 til 6 ), brugte vi en generaliseret lineær blandet model (GLMM), der inkluderede disse fem forudsigere som faste effekter og udløsere (Trigger) og respondenter (Responder) identiteter som tilfældige effekter. Vi monterede modellerne i R (R Core Team, 2018; version 3.5.1) ved hjælp af funktionen lmer af R-pakken lme4 (Bates et al., 2015). Vi fastslog betydningen af den fulde model ved sammenligning med en nulmodel, der kun omfatter tilfældige effekter (Forstmeier og Schielzeth, 2011). Vi brugte en sandsynlighedsforholdstest (Dobson, 2002) for at teste denne betydning (ANOVA med argumentet “Chisq”). Vi beregnede p-værdierne for de enkelte forudsigere baseret på sandsynlighedsforholdstest mellem den fulde og nulmodellen ved hjælp af R -funktion “drop1” (Barr et al., 2013). Da svarvariablen var binær, brugte vi en binomial fejlfordeling. Vi testede om interaktionen mellem kønnene eller triggerklassens aldersklasser og responderen var signifikant, men da de ikke var, inkluderede vi dem ikke i modellen. Vi brugte en multipel kontrastpakke (multcomp) til at udføre alle parvise sammenligninger for hvert bindingsniveau med Tukey-testen (Bretz et al., 2010). Vi rapporterede de Bonferroni-justerede p-værdier, estimat (Est), standardfejl (SE) og z-værdier.
Resultater

Figur 1. Linjeplot af virkningen af det sociale bånd mellem trigger og responder (X-akse) på den gennemsnitlige forekomst af akustisk gabe-smitte (Y-akse). Venner og pårørende viser signifikant højere gaben smittefrekvenser end fremmede og bekendte (Tukey test: venner versus fremmede p < 0.001; slægtning mod fremmede p < 0,001; venner versus bekendte p < 0,001; familie versus bekendte p < 0,001; andre kombinationer, ns).
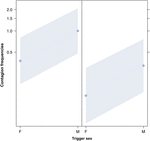
Figur 2. Linjeplot af effekten af udløserens køn (X-aksen) på den akustiske gabe-smitte middel forekomst (Y-aksen). Effekt af trigger-køn på akustisk gaben smitte, når responderen er en kvinde (højre) og en mand (venstre). Hannes gab fremkalder flere gaben end hunnernes uanset køn af responderen (resultat af GLMM, p = 0,022). Point repræsenterer den effekt, som responsvariablen har på den uafhængige variabel “trigger of sex”, baseret på den forudsagte værdi af modellen. Farvede bånd viser 95% konfidensinterval.
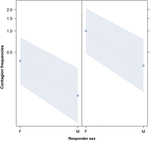
Figur 3. Linjeplot af effekten af køn af responder (X-akse) på akustisk gabe-smitte middel forekomst (Y-akse). Effekt af responder-køn på akustisk gabe-smitte, når udløseren er en kvinde (højre) eller en mand (venstre). Kvinder reagerer markant mere end mænd uanset af udløserens køn (resultat af GLMM, p = 0,021). Punkter repræsenterer den effekt, som responsvariablen har på den uafhængige variabel “responderens køn”, baseret på den værdi, der er forudsagt af modellen. Farvede bånd viser 95% konfidensinterval.
Diskussion
Denne undersøgelse viser for første gang, at gabesmitte er væsentligt påvirket af den sociale bånd mellem individer (tabel 1), selv når de udløsende stimuli er auditive gab, som vi definerede som vokaliserede gabber, der kunne høres, men ikke ses (visuel cue, der ikke kan detekteres, auditiv cue detekterbar). Især er auditiv smitsom gaben signifikant hyppigere mellem slægtninge og venner end mellem fremmede og bekendte (figur 1). Dette fund understøtter forudsigelse 1a baseret på EBH og ikke forudsigelse 1b baseret på ABH, hvilket fører til den konklusion, at selektiv visuel opmærksomhed ikke kan være den vigtigste drivkraft for den sociale asymmetri hos mennesker, der observeres i gabesmittehastigheder (Norscia og Palagi). , 2011; Norscia et al., 2016a). Også udløserens køn og modtagerens køn havde en signifikant effekt på gabesmittehastigheder, hvor mænd – som udløsere – blev reageret hyppigere af andre end kvinder (figur 2), og kvinder reagerede oftere på andres gabe mænd (figur 3).
I modsætning til Bartholomew og Cirulli (2014) fandt vi ingen alderseffekt på gaben smitte, sandsynligvis fordi vores database om hørselsgabe havde en stærk forekomst af voksne (25-64 år gammel). De højeste niveauer af auditiv gaben smitte hos kvinder sammenlignet med mænd bekræfter kønsforstyrrelse observeret under naturalistiske forhold hos mennesker, der er modtagelige for gaben smitte af Norscia et al. (2016a, b) når man overvejer et større datasæt, der også omfattede gaben, der kunne ses af den potentielle responder (med gaben sensorisk modalitet – syn, hørelse eller begge dele – som ikke påvirker responsen). Kønsforstyrrelsen er også delvis enig med tidligere resultater opnået i kontrollerede indstillinger, herunder den visuelle signal (Chan og Tseng, 2017; men se Norscia og Palagi, 2011; Bartholomew og Cirulli, 2014). Det er blevet antaget, at den høje grad af gaben smitte hos kvinder kan informere følelsesmæssig smitte (Norscia et al., 2016a) i lyset af kvindernes højere empatiske evner – relateret til moderskab – sammenlignet med mænd (Christov-Moore et al. ., 2014). Imidlertid er dette spørgsmål stadig under debat, fordi kulturelle forskelle på tværs af menneskelige samfund kan forme den sociale bindingsdynamik på en anden måde. Det er derfor kompliceret på dette stadie af viden at skille kulturelle faktorer, interpersonel forholdskvalitet og kønsindflydelse fra hinanden i fordelingen af gaben smitte. Vores resultater viser også, at mænd fungerer bedre end kvinder som udløsere, og den mest parsimoniske hypotese for dette kan være, at mænds vokaliseringer kan høres bedre i naturlige omgivelser, ofte præget af baggrundsstøj. Faktisk er opfattelsen af stemmekøn primært afhængig af den grundlæggende frekvens, der i gennemsnit er lavere med en oktav hos mandlige stemmer end kvindelige stemmer, hvor vokaliseringer med lavere frekvens bevæger sig længere end højfrekvente dem (Marten og Marler, 1977; Latinus og Taylor, 2012) . Så vidt vi ved, er der ingen specifik undersøgelse, der adresserer den mulige kønsforstyrrelse i gabenhørbarhed, og yderligere undersøgelse med eksperimentelle forsøg i kontrolleret tilstand er derfor nødvendig for at bekræfte denne spekulation.
I denne undersøgelse fandt vi også at forskellene i gabesmittehastigheder på tværs af kategorier (familie og venner, fremmede og bekendte) ikke kan forklares med forskelle i top-down, selektiv visuel opmærksomhed. Dette fund er i tråd med tidligere litteratur. Smitsom gaben synes at involvere hjerneområder, der er mere relateret til det orienterende bund-op-netværk end top-down-relaterede områder. Desuden er gaben smitte hverken følsom over for de sensoriske signaler, der er til stede i signalet (auditive, visuelle eller audiovisuelle) (Arnott et al., 2009; Norscia og Palagi, 2011) eller påvirkes af det visuelle perspektiv af den udløsende stimulus ( gab i orienteringer på 90 °, 180 ° og 270 ° er i stand til at udløse gabresponser som frontal, 0 ° gab; Provine, 1989, 1996). Chan og Tseng (2017) fandt, at evnen til at detektere et gaben som sådan (perceptuel detektionsfølsomhed) var relateret til varigheden af blik til øjnene på det stimulus, der frigør ansigt, men øjenblikkemønstre var ikke i stand til at modulere smitsom gab. I chimpanser var smitsomme gabefrekvenser højest mellem samme gruppe end individer i forskellige grupper, selvom respondenterne kiggede længere på chimpansevideoer uden for gruppen (Campbell og de Waal, 2011). Argumentet om, at visuel selektiv opmærksomhed kan forspænde gabe smittehastigheder i en bestemt retning (forsøgspersoner, der reagerer mere på familie end fremmede) undermineres også af fraværet af noget specifikt mønster for social opmærksomhed hos menneskelige og ikke-menneskelige primater. Via eye-tracking (anvendt til at måle visningstiden) og ved at vise ukendte ansigter til deres eksperimentelle emner, Méary et al.(2014) observerede, at mennesker var skæv mod ansigt fra egen race, mens rhesusmakakernes opmærksomhed blev mere tiltrukket af nye end af samme artsansigter. Kawakami et al. (2014) observerede, at mennesker blev mere opmærksomme på øjnene på etniske medlemmer i gruppen og på næsen og munden på etniske medlemmer af gruppen. Den samme undersøgelse afslørede også, at visuel opmærksomhed ikke var afhængig af målløb. Ved at måle, hvor længe de forsøgspersoner så på skærmen, Whitehouse et al. (2016) bemærkede, at Barbary-makakerne var mere opmærksomme på at skrabe videoer af ikke-fremmede end fremmede individer, men bemærkede også, at inden for ikke-fremmede var makakker mest opmærksomme på de personer, som de delte en svag social binding med. Ved at måle blikfrekvenser observerede Schino og Sciarretta (2016), at mandrills så mere på deres egne slægtninge end på ikke-pårørende, men også mere på dominerende end hos underordnede gruppekammerater. Derfor beskriver disse undersøgelser (brugt til understøttelse af ABH) intet enkelt mønster af selektiv opmærksomhed. Et yderligere vigtigt punkt at overveje er selve definitionen af fortrolighed og gruppemedlemskab, der er vedtaget af de fleste af de undersøgelser, der blev brugt til at støtte ABH (Massen og Gallup, 2017). Disse undersøgelser viste bedre visuel påvisning og visuel perceptiv kodning af ansigter til velkendte / i-gruppe forsøgspersoner sammenlignet med ukendte, men definerede fortrolighed og gruppemedlemskab ikke på baggrund af de personlige forhold mellem individer. I stedet blev fortrolighed eller gruppemedlemskab defineret på grund af indirekte viden (fx foto af berømte personer eller af et emne, der allerede er vist i fasen forud for retssagen) eller fælles race (f.eks. Buttle og Raymond, 2003; Ganel og Goshen- Gottstein, 2004; Jackson og Raymond, 2006; Michel et al., 2006). Denne definition er fint i forbindelse med disse undersøgelser, men det er ikke så meget fint, hvis resultaterne bruges til at foreslå alternative forklaringer på den indflydelse, som reel social binding – baseret på reelle forhold – kan have på et fænomen, i dette tilfælde gabe smitte . For eksempel Michel et al. (2006) observerede, at kaukasiske og asiatiske forsøgspersoner bedre kunne genkende ansigter af samme race, men denne forskel var ikke til stede hos asiatiske forsøgspersoner, der havde boet i omkring et år blandt kaukasiere. Et andet diskussionspunkt vedrører tilstedeværelsen af gaben smitte hos børn med autismespektrumforstyrrelse (ASD), som ofte viser ændringer i visuel opmærksomhed (Richard og Lajiness-ONeill, 2015). Hos ASD-børn kan gabesmitte være fraværende (Senju et al., 2007), nedsat (Helt et al., 2010) eller lignende til typisk udviklende børn, når forsøgspersonerne induceres til at omdirigere deres opmærksomhed videostimulus under de eksperimentelle forsøg (Usui et al., 2013). I en nylig undersøgelse har Mariscal et al. (2019) fandt ud af, at gabenesmitte hos ASD-børn var positivt relateret til blodkoncentrationen af oxytocin, hormonet involveret i forældrenes og sociale tilknytning (Decety et al., 2016) og hævdede, at gabenesmitte hos ASD-børn kan være relateret til variabelt gennemsnit oxytocinkoncentrationer på tværs af forskellige undersøgelseskohorter (Mariscal et al., 2019). Dette fund er i tråd med EBH-hypotesen, der forbinder gabe-smittehastigheder med social binding, hvilket kan afspejle følelsesmæssig binding.
Konklusion
Vores undersøgelse føjer til diskussionen om de mekanismer, der ligger til grund for social asymmetri i gaben smitte (for kritiske anmeldelser: se Adriaense et al., 2020; Palagi et al., 2020) ved at vise, at gaben smitte sandsynligvis er forbundet med nedenfra og op snarere end med top-down, selektiv opmærksomhed. Bottom-up-opmærksomhed ledes primært af den sensoriske opfattelse af den fremkaldende stimulus, hvorimod selektiv opmærksomhed fra oven er en frivillig, vedvarende proces, hvor en bestemt genstand vælges internt og fokuseres på eller undersøges (Katsuki og Constantinidis, 2014). I denne henseende blev den akustiske stimulus (auditiv gab), der blev udsendt af aftrækkeren, hørt og kunne fremkalde et gabende svar i modtageren, selvom modtageren ikke betød nogen frivillig visuel opmærksomhed på udløseren. Desuden blev de gabende responsrater socialt moduleret, idet auditiv gaben smitte var højest hos personer, der var stærkest bundet til hinanden. Derfor er selektiv opmærksomhed fra oven ikke den vigtigste drivkraft for den sociale asymmetri, der observeres i gaben smitte, hvilket ser ud til at være et stimuleringsdrevet fænomen relateret til nedenfra og op opmærksomhedsprocesser. Yderligere undersøgelse er nødvendig for at forstå, om og på hvilken måde andre former for opmærksomhed eller præ-opmærksomme stadier er i stand til at påvirke gaben smitte.
Erklæring om datatilgængelighed
Datasættet, der blev brugt til denne undersøgelse findes i det supplerende materiale.
Etisk erklæring
Undersøgelsen, der involverede menneskelige deltagere, blev gennemgået og godkendt af Comitato di Bioetica dAteneo (University Bioethical Committee) —University of Turin (ref. nr. 451945).Skriftligt informeret samtykke til deltagelse var ikke påkrævet for denne undersøgelse i overensstemmelse med den nationale lovgivning og de institutionelle krav.
Forfatterbidrag
IN og EP gennemførte dataindsamling og udtænkte og skrev manuskript. AZ hjalp med dataindsamling og manuskriptrevision, figurer og tabeller. MG udførte statistiske analyser og skrev den relaterede del af manuskriptet.
Interessekonflikt
Forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Anerkendelser
Forfatterne ønsker at takke prof. Roberto Barbuti og prof. Cristina Giacoma for at have støttet forskningen ved universitetet i Pisa og Universitetet i Torino henholdsvis.
Supplerende materiale
Altmann, J. (1974). Observationsundersøgelse af adfærd: prøvetagningsmetoder. Opførsel 49, 227-226. doi: 10.1163 / 156853974×00534
PubMed Abstract | CrossRef fuldtekst | Google Scholar
Anderson, J. R. og Meno, P. (2003). Psykologiske påvirkninger på gaben hos børn. Curr. Psykol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J. og Cirulli, E. T. (2014). Individuel variation i smitsom følsomhed over for gaben er meget stabil og stort set uforklarlig af empati eller andre kendte faktorer. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | CrossRef fuldtekst | Google Scholar
Bretz, F., Hothorn, T. og Westfall, P. (2010). Flere sammenligninger ved hjælp af R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Etologisk undersøgelse af gaben hos primater. I. Kvantitativ analyse og undersøgelse af årsagssammenhæng hos to arter af gamle verdensaber (Cercocebus albigena og Macaca fascicularis). Etologi 98, 221-245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Fuld tekst | Google Scholar
Dobson, A. J. (2002). En introduktion til generaliserede lineære modeller, 2. udgave. Boca Raton, FL: Chapman og Hall.
Google Scholar
Katsuki, F. og Constantinidis, C . (2014). Bottom-up og top-down opmærksomhed: forskellige processer og overlappende neurale systemer. Neurovidenskab 20, 509-521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef fuldtekst | Google Scholar
Singer, T. (2006). Det neuronale grundlag og ontogeni af empati og tankelæsning: gennemgang af litteratur og implikationer for fremtidig forskning. Neurosci. Biobehav. Rev. 30, 855-863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | CrossRef fuldtekst | Google Scholar
Statistics Canada (2009). Alderskategorier, grupperinger af livscyklus. Ottawa: Statistics Canada.
Google Scholar