Zpracování informací je nezbytné ve všech vědních oborech. V molekulární biologii je centrální dogma, které poprvé vytvořil Francis Crick (Crick, 1958, 1970), klasickou páteří živých buněk k zásadnímu provádění procesů od dělení buněk po smrt prostřednictvím informačních cest DNA, RNA a bílkovin. Přesněji řečeno, centrální dogma popisuje přenos informací o sekvenci během replikace DNA, transkripci do RNA a translaci do aminokyselinových řetězců tvořících proteiny. Současně také uvádí, že informace nemohou proudit z proteinu na protein nebo nukleovou kyselinu.
Od nástupu systémových a vysokovýkonných přístupů v posledních dvou desetiletích tyto široké kroky, které nezahrnují složité regulační podrobnosti, byly podrobeny intenzivní kontrole. Chybějící regulační prvky, jako jsou mechanismy korektury / opravy DNA a alternativní sestřih pre-mRNA, zavádějí několik zprostředkujících kroků. Tyto další kroky zasahují do klíčových kroků dogmatu a pravděpodobně mění dynamiku informací. Navíc se zdá, že epigenetika neboli role chromatinových struktur, methylace DNA a modifikace histonů je v rozporu s jednoduchými cestami dogmatu (Shapiro, 2009; Luco et al., 2011). Spojování proteinů nebo schopnost proteinu (inteiny) měnit svou vlastní sekvenci objevenou v poslední době (Volkmann a Mootz, 2012) a priony, které modifikují další proteinové sekvence (Prusiner, 1998), obcházejí cestu přenosu informací dogma. Další výzkumy uváděly chyby nebo neshody mezi sekvencemi RNA a jejich kódující DNA (Hayden, 2011; Li et al., 2011). Dohromady tyto údaje zpochybňují platnost centrálního dogmatu v kontextu současné vědy, a proto zpochybňují jednoduchost lineárního toku informací (DNA k RNA a RNA k proteinu).
Abychom věci uvedli na pravou míru, požadujeme analytické nástroje, které zkoumají obavy nebo nesrovnalosti týkající se dlouhodobé teorie. Jednou z jednoduchých, ale velmi užitečných technik pro vyhledávání globálních vlastností ve vysoce výkonných souborech dat je statistická korelační analýza, která se široce a úspěšně používá k pozorování vzorů ve složitých systémech, jako je počasí (Stewart, 1990), akciové trhy (Lo a MacKinlay , 1988) a kosmologie (Amati et al., 2008). Existuje několik druhů korelačních analýz, které hodnotí jak lineární (např. Pearsonův produktový moment), tak nelineární (např. Spearmanova hodnost, vzájemné informace) závislosti (Steuer et al., 2002; Rosner, 2011). Zejména , Pearsonova produktová momentová korelační analýza se stala nejpopulárnější díky své schopnosti ukázat organizační strukturu v nejjednodušší formě.
V biologii existuje řada prací, které se zabývaly korelacemi v mRNA a data exprese proteinů (viz níže a tabulka 1). Teoreticky, když jsou porovnány dva vzorky obsahující vysoce dimenzionální (jako jsou mikročipová a proteomická) data, poskytují korelační analýzy míru odchylky od jednoty jako zdroje rozdílu mezi vzorky . Stručně, dva vzorky se identickými a zcela neidentickými informacemi ukážou jednotkovou (R2 = 1) a nulovou (R2 = 0) korelaci.
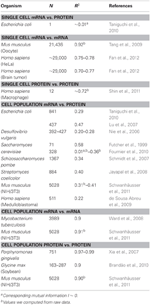
Tabulka 1. mRNA a protein korelace relace v různých organismech.
Dokonalá korelace (R2 = 1) je idealizovaná situace, která je daleko od reality, protože je technická nebo experimentální šum sám interferuje a snižuje korelaci. Poslední roky navíc zdůraznily existenci biologického šumu: studie na jednotlivých buňkách a molekulách ukázaly stochasticitu v dynamice genové exprese díky kombinatorickému účinku nízkého počtu molekulárních kopií a kvantové povaze dynamiky promotorů (Raj a van Oudenaarden, 2009; Eldar a Elowitz, 2010). Na druhé straně klonální populace buněk vykazují heterogenitu v úrovních dané exprese proteinu na buňku v jakémkoli měřeném čase (Chang et al., 2008). Společně je stochasticita a heterogenita nezbytná pro produkci diverzifikace osudu buněk, fenotypových variací a zesílení intracelulárních signálů (Locke et al., 2011; Selvarajoo, 2012).
Stochastické fluktuace neboli vnitřní šum způsobují exprese molekulárních druhů se mění v čase a mezi buňkami, což vede k nekorelovaným reakcím (Elowitz et al., 2002). To je zvláště prominentní pro mRNA a proteiny s nízkým počtem kopií. Korelaci mezi vzorky (buňkami) lze tedy snížit kvůli vnitřnímu šumu (obrázek 1A). Mezi další zdroje biologického šumu způsobeného vnějšími faktory patří variabilita velikosti buněk, počet molekulárních kopií a výkyvy prostředí mezi jednotlivými buňkami.Tyto faktory narušují deterministické centrální dogma a pravděpodobně mění silné korelace na slabší (obrázek 1B).
Jedna nedávná studie srovnávala expresi mRNA a proteinů Escherichia coli mezi jednotlivými buňkami na úrovni jedné molekuly a poskytla scénář, který hluboce zpochybňuje centrální dogma. Taniguchi a kol. (2010) odhalili, že neexistuje žádná korelace (R2 ~ 0) mezi jednotlivými tufA mRNA a hladinami proteinů v jednotlivých buňkách. Zejména dospěli k závěru, že nedostatek korelace je pravděpodobně způsoben rozdíly v životnosti mRNA a bílkovin. Přestože se jedná o věrohodné vysvětlení, Taniguchi a kol. byli opatrní, aby nevyvrátili dlouhodobou hypotézu tvrzením, že časové průměry hladin mRNA by měly korelovat s hladinami proteinů. Nebyl však prokázán žádný důkaz, který by prokázal, že se jedná o skutečný případ, a když jsme hodnotili nelineární závislosti pomocí vzájemných informací (Steuer et al., 2002; Tsuchiya et al., 2010) v Taniguchi et al. datová sada, zjistili jsme, že výsledek není závislý, tj. I ~ 0. To potvrzuje, že exprese mRNA na protein mezi jednotlivými buňkami na úrovni jedné molekuly zjevně nesouvisí. Při zvětšování na úrovni jedné molekuly v korelačním grafu je dále zřejmé, že jejich párové korelace jsou slabé (obrázek 1A, pro ilustraci).
Zejména na úrovni buněčné populace Taniguchi et al. byli schopni ukázat relativně vysokou korelaci mezi expresí mRNA a proteinu s R2 = 0,29 (obrázek 2A). Ve skutečnosti další nezávislá studie Lu et al. (2007), pro populaci E. coli, také vykázal relativně vysokou korelaci (R2 = 0,47). Podobné analýzy prováděné na Saccharomyces cerevisiae (Futcher et al., 1999), myších fibroblastech NIH / 3T3 (Schwanhäusser et al., 2011) a několika dalších populacích buněk (Nie et al., 2006; Schmidt et al., 2007; Jayapal et. al., 2008; de Sousa Abreu et al., 2009) všechny vykazovaly korelované struktury mezi expresemi v celém transkriptomu a v proteomu (tabulka 1). Proč tedy neexistuje korelace mezi expresí jednotlivých mRNA a proteinů v jednotlivých buňkách, zatímco na úrovni populace jsou pozorovány kolektivní vztahy mezi expresemi mRNA a proteinů ve velkém měřítku?
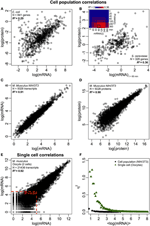
Obrázek 2. Korelace výrazů v celém Omics. Populace buněk: korelace mRNA-protein v (A) E. coli (Taniguchi et al., 2010) a (B) S. cerevisiae (Fournier et al., 2010) mezi expresemi mRNA v čase t = 60 minut a proteinovými expresemi v čase t = 360 min. Vložit: korelační matice mezi všemi časovými body ukazuje opožděný nárůst korelací mezi mRNA a proteiny. (C) mRNA a (D) proteinové exprese mezi dvěma vzorky myších buněk NIH / 3T3 (Schwanhäusser et al., 2011). Jednotlivé buňky: (E) exprese mRNA mezi dvěma oocyty (Tang et al., 2009). Červené tečkované čáry označují oblasti s nízkou expresí mRNA (log (mRNA) < 5). (F) Šum (η2) versus log (mRNA exprese) pro buněčnou populaci (NIH / 3T3, černé tečky, Schwanhäusser et al., 2011) a jednotlivé buňky (Oocyty, zelené trojúhelníky, Tang et al., 2009). Každá tečka představuje hodnotu pro skupinu P = 100 mRNA. η2 je téměř nulová pro buněčnou populaci pro všechny výrazy mRNA. U jednotlivých buněk je η2 nejvyšší pro mRNA s nejnižším počtem kopií a blíží se nule pro vyšší počty kopií.
Věříme existují dva hlavní důvody rozdílů. Za prvé, jak již bylo uvedeno dříve, hluk, ať už biologické nebo nebiologické povahy, snižuje korelaci. Vzhledem k tomu, že analýzy na jednotlivých buňkách prokázaly význam stochasticity a variability, jsou tyto účinky zásadní pro snížení korelace jednotlivých buněk. Na úrovni souboru, když jsou buňky vzorkovány do populace, je celkový (vnitřní + vnější) hluk snížen, protože náhodný hluk se ruší v celém rozsahu molekulárních výrazů (obrázky 1C – F), aby odhalil průměrnou odezvu a sebeorganizaci ( Karsenti, 2008; Selvarajoo, 2011; Hekstra a Leibler, 2012; Selvarajoo a Giuliani, 2012). Proto se objevuje dobrý stupeň korelace exprese mRNA-protein. Za druhé, pro studii s jednou buňkou (Taniguchi et al., 2010) byla porovnána korelace exprese jednotlivých mRNA-protein napříč mnoha buňkami. Ve studiích buněčné populace se však provádí srovnání jako celek, přes tisíce mRNA a proteinů o několik řádů větších, než je rozsah exprese nalezený pro jednu molekulu mezi buňkami. To proto vede k vyšším korelacím na úrovni populace, protože účinek jednotlivých molekulárních variací je zanedbatelný.
Navzdory pozorovaným korelovaným strukturám u buněčných populací existují hmatatelné důvody pro velkou odchylku od dokonalé korelace.Jak již bylo uvedeno dříve, jedním klíčovým bodem je, že mRNA a proteiny jsou postupně lokalizovány s několika chybějícími procesy, které nejsou zastoupeny v centrálním dogmatu. Přidání chybějících meziproduktů podél biochemické cesty způsobí znatelné zpoždění v toku informací (Selvarajoo, 2006, 2011; Piras et al., 2011) a výsledkem by mohla být korelace mezi nimi. To by mohlo být také součástí skutečnosti, kterou zaznamenali Taniguchi a kol. že exprese mRNA a proteinu mají různou životnost. Je pozoruhodné, že tato postulace je podporována v nedávné práci na S. cerevisiae léčené Rapamycinem, která ukázala, že časové korelace exprese mRNA-proteinu byly zpočátku nízké, R2 = 0,01 ve 40 minutách, nicméně během 360 minut po poruše se korelace zvýšila, R2 = 0,36 (Fournier et al., 2010, obrázek 2B). Data naznačují, že při chemické perturbaci se počáteční reakce mezi expresí mRNA a proteinu odchyluje v důsledku časového zpoždění a různých kinetických mechanismů mezi nimi, jakož i sekundárních účinků, jako je autokrinní nebo parakrinní signální interference (Shvartsman et al., 2002; Isalan et al., 2008). Když jsou účinky narušení v průběhu času zeslabeny, došlo k obnovení korelací.
Abychom dále ověřili postulaci, že pro snížení korelací mRNA-protein jsou rozhodující procesy sekvenčního zpoždění nebo různé doby života, porovnali jsme R2 mezi stejné molekulární druhy centrálního dogmatu (např. mezi mRNA a mRNA) v buněčných populacích a jednotlivých buňkách. Transkriptomová korelace mRNA-mRNA exprese mezi replikáty NIH / 3T3 (Schwanhäusser et al., 2011) (obrázek 2C) a vzorky buněčné populace Mycobacterium tuberculosis (Ward et al., 2008) jsou oba velmi vysoké, s R2 > 0,9 (tabulka 1). Takové silné korelace lze pozorovat také mezi populačními vzorky pro expresi protein-protein v buňkách NIH / 3T3 (Schwanhäusser et al., 2011) (obrázek 2D), Porphyromonas gingivalis (Xia et al., 2007) a Glycine max (Brandão et al. , 2010) (tabulka 1). Protože tato data, která srovnávají stejné druhy, poskytují velmi vysoké korelace, je možné, že procesy postupného zpoždění nebo různé doby života jsou odpovědné za snížení korelačních struktur na úrovni populace mezi expresí mRNA a proteinů.
V jednotlivých myších oocytech ( Tang et al., 2009), při srovnání celých expresí mRNA – mRNA je pozorována vysoce korelovaná struktura (R2 = 0,92, obrázek 2E). Soustředíme-li se však pouze na málo exprimované mRNA (s logaritmickými výrazy < 5), stochastický šum poměrně výrazně snižuje párovou korelaci (R2 < 0,54). K otestování tohoto výsledku jsme vyhodnotili šum, η2 = σ2XY / μ2XY, napříč celými výrazy mRNA (obrázek 2F). Poznamenali jsme, že η2 je nejvyšší pro nejnižší výrazy, kvůli výraznému účinku stochastických fluktuací ve srovnání s jejich výrazy, a blíží se nule pro vyšší výrazy, kde se takový šum stává méně významným (Piras et al., 2012). U buněčné populace, jak se očekávalo, je pozorován téměř nulový šum v celém rozsahu exprese kvůli rušení náhodného šumu (obrázky 1E, F).
Rovněž byly vysoce korelované struktury pro celou expresi mRNA – mRNA hlášeny pro jednotlivé rakovinné buňky (Fan et al., 2012), i když méně významné s R2 ~ 0,7 (tabulka 1). Kromě toho srovnání expresí proteinů a proteinů v lidských makrofágech stimulovaných LPS také ukázalo vysoké korelace, R2 ~ 0,72 (Shin et al., 2011) (tabulka 1). Ačkoli neexistuje žádná korelace mezi jednotlivými expresemi mRNA-proteinu v jednotlivých buňkách, je korelace ve velkém měřítku nebo v širokém rozsahu mezi stejnými molekulárními druhy v jednotlivých buňkách velmi vysoká.
Tedy, ať už jednotlivé buňky nebo buněčné populace , údaje z celého omik naznačují, že korelace mezi stejnými molekulárními druhy (mRNA vs. mRNA a protein vs. protein) jsou znatelně vyšší než mezi různými druhy (mRNA vs. protein). To odráží skutečnost, že ačkoliv jsou procesy zpoždění a různé doby života klíčem ke snížení korelací, tyto mechanismy nejsou dostatečné pro podporu chybějící korelační struktury pozorované mezi jednotlivými transkripcemi jednotlivých proteinů na expresi jednotlivých buněk.
Takže daleko, zkoumáním rozsáhlých expresí mRNA a proteinů různých buněčných systémů, jsme ukázali, že korelační struktury vznikají v globálním měřítku. korelační analýzy však odhalují pouze konektivitu mezi dvěma testovanými vzorky a neukazují směr tok informací. Aby centrální dogma platilo v globálním měřítku, měl by být celkový tok informací od DNA k proteinům. Tento tok informací byl prokázán nesčetnými dalšími studiemi, které zahrnují narušení receptorů buněčných populací a sledování výsledného dynamika transkripčních faktorů vázajících se na DNA a indukce genových expresí ve velkém měřítku (obrázek 3A).Například v případě LPS-stimulovaných imunitních buněk bylo prokázáno, že k aktivaci transkripčního faktoru NF-kB dochází přibližně za 15 minut (Liu et al., 1999), indukce jeho následných genů přibližně 30 min (Liu et al., 1999; Xaus et al., 2000; Selvarajoo et al., 2008) a translace odpovídajících proteinů v oblasti 60–90 min (Kawai et al., 1999; Xaus et al. ., 2000) (Obrázek 3B). Takový postupný směr celkového toku transkripce do toku translačních informací lze pozorovat také u bakteriálních systémů, jako je E. coli, na úrovni buněčné populace (Golding et al., 2005).

Obrázek 3. Tok informací centrálního dogmatu. (A) Schéma exprese TNF indukované LPS / TLR4 prostřednictvím transkripčního faktoru NF-kB a genu tnf, po lineárním toku informací. (B) Experimentální časové profily vazebné aktivity promotoru expresí NF-kB (horní panely), tnf (střední panely) a TNF (spodní panely) na úrovni buněčné populace. (C) Schematické časové profily dynamiky promotorů, mRNA a expresí proteinů na úrovni jednotlivých buněk (Raj a van Oudenaarden, 2009).
Alternativně výzkumy v rozlišení jednotlivých buněk odhalily náhodné výkyvy lineárního toku informací: transkripční faktory vázající se na oblasti promotoru DNA jsou kvantové, což má za následek praskavé chování transkripce mRNA a následně indukuje variabilitu v translaci proteinu, dokonce mezi identickými buňkami (obrázek 3C) (Raj a van Oudenaarden, 2009; Eldar a Elowitz, 2010; Locke et al., 2011; Hekstra a Leibler, 2012; Selvarajoo, 2012). Výsledkem je, že v kterémkoli konkrétním časovém bodě je individuální molekulární odezva pro jednotlivé buňky ve srovnání s průměrnou populační škálou poměrně hlučná (Selvarajoo, 2011).
Závěry
Příklady uvedené v tento článek zdůrazňuje rozdíly v pořadí hodnot korelace pozorovaných mezi druhy v centrálním dogmatu nad buněčnými populacemi a jednotlivými buňkami. Statistické analýzy z buněčných populací vykreslují obraz, že korelace exprese mezi stejnými molekulárními druhy je velmi vysoká a mezi druhy je mírně vysoká. I když jsou korelace jednotlivých buněk mezi stejnými druhy srovnatelné s buněčnými populacemi, vykazovaly širší rozptyl v grafech jejich expresí kvůli výraznému účinku biologického šumu, zejména u transkriptů s nízkým počtem kopií. Je pozoruhodné, že párová korelace jednotlivých buněk se pro jednotlivé molekuly stává nulovou (Taniguchi et al., 2010). Ve skutečnosti je známo, že stochastické fluktuace a variabilita v molekulárních expresích jsou funkční při generování rozhodování o osudu buňky a tipování buněčných stavů (Losick a Desplan, 2008; Eldar a Elowitz, 2010; Kuwahara a Schwartz, 2012). Věříme, že k silným korelacím v celé oblasti dochází v důsledku těsných regulačních sítí genů a proteinů napříč tisíci molekul (Barabási a Oltvai, 2004; Karsenti , 2008), což má za následek vznikající průměrné odpovědi. Při analýze malého počtu nebo jednotlivých molekul nelze korelační strukturu pozorovat.
Celkově je možné, že prohlížení informačního toku jedné DNA k proteinu zpochybní centrální dogma protože odpověď každé molekuly v kterémkoli okamžiku pravděpodobně nebude korelovat. Z globálního hlediska však pozorování průměrné deterministické odpovědi naznačuje, že čistá rovnováha Úplně vpravo od cest zůstávají ic informace. Centrální dogma by proto mělo být vnímáno jako makroskopický buněčný informační tok v širokém měřítku, nikoli na úrovni jednoho genu a proteinu. Proto věříme, že jeho jednoduchost zůstane jedním z nejvlivnějších teoretických pilířů živých systémů.
Konflikt zájmů
Autoři prohlašují, že výzkum byl proveden v absence jakýchkoli obchodních nebo finančních vztahů, které by mohly být považovány za potenciální střet zájmů.
Poděkování
Kentaro Hayashi se děkuje za komentáře. Oceňuje výzkumný fond města Tsuruoka a prefektury Yamagata.
Crick, F. (1958). Na syntézu bílkovin. Symp. Soc. Exp. Biol. 12, 139–163.
Pubmed Abstract | Plný text Pubmed
Crick, F. (1970). Centrální dogma molekulární biologie. Nature 227, 561–563.
Pubmed Abstract | Celý text Pubmed
Hayden, E. C. (2011). Důkazy o změněné RNA vyvolávají debatu. Nature 473, 432.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Hekstra, D. R. a Leibler, S. (2012). Pohotovostní a statistické zákony v replikovaných mikrobiálních uzavřených ekosystémech. Buňka 149, 1164–1173.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Kuwahara, H. a Schwartz, R. (2012).Zisk stochastického ustáleného stavu v procesu genové exprese s kontrolou degradace mRNA. J. R. Soc. Rozhraní 9, 1589–1598.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Nie, L., Wu, G. a Zhang, W. (2006). Korelace exprese mRNA a nadbytku bílkovin ovlivněna mnoha sekvenčními rysy souvisejícími s translační účinností u Desulfovibrio vulgaris: kvantitativní analýza. Genetics 174, 2229–2243.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Prusiner, S. B. (1998). Priony. Proc. Natl. Acad. Sci. USA 95, 13363–13383.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Rosner, B. (2011). Základy biostatistiky. 7. vydání Boston, MA: Duxbury Press.
Selvarajoo, K. (2006). Objevení mechanismů diferenciální aktivace signálních drah podobného mýtnému receptoru 4 v knockoutech MyD88. FEBS Lett. 580, 1457–1464.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Selvarajoo, K. (2011). Makroskopický zákon zachování odhalený v populační dynamice signalizace receptoru podobného mýtnému. Cell Commun. Signál. 9, 9.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Selvarajoo, K. (2012). Pochopení multimodálních biologických rozhodnutí z dynamiky jedné buňky a populace. Wiley Interdiscip. Rev.Syst. Biol. Med. 4, 385–399.
Pubmed Abstract | Plný text Pubmed | CrossRef Full Text
Stewart, T. R. (1990). Rozklad korelačního koeficientu a jeho použití při analýze prognostických dovedností. Předpověď počasí. 5, 661–666.