Úvod
Zívání je nedobrovolná sekvence otevírání úst, hluboká inspirace, krátká apnoe a více či méně pomalé výdechy (Baenninger, 1997; Walusinski a Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Při vyvolání nelze zívnutí zcela potlačit. Proto byl definován jako stereotypní nebo reflexní vzor (Lehmann, 1979; Provine, 1986). V Homo sapiens bylo navrženo několik hypotéz s různou podporou vysvětlujících mechanismy a funkce spontánního zívání, jako je okysličování (funkce dýchání způsobená hypoxií), chování související se stresem (způsobené vzrušením) nebo termoregulace (způsobená hypertermií; Guggisberg et al., 2010; Massen et al., 2014; Gallup a Gallup, 2019). Jako fyziologická reakce může být zívání ovlivněno vnitřními a vnějšími faktory, jako je denní doba (Giganti a Zilli, 2011) nebo intrakraniální / mozková teplota (Gallup a Eldakar, 2013).
Zívání může být sebeřízený a / nebo zobrazen ostatním (Moyaho et al., 2017; Palagi et al., 2019). U lidských a subhumánních primátů může v závislosti na druhu, když je zívání ostatním prokázáno, komunikovat ohrožení (Troisi et al., 1990; Deputte, 1994) a / nebo fyziologické a behaviorální změny (Provine et al., 1987) ; Leone et al., 2015; Zannella et al., 2015). U lidí je zívání sociálně modulovanou odpovědí, protože může být potlačeno skutečnou – a nikoli virtuální – sociální přítomností (Gallup et al., 2019) a protože zívání může být vyvoláno zíváním někoho jiného v důsledku známého jevu jako nakažlivé zívání (Provine, 1989, 2005). Nákazu zíváním lze vyvolat, i když je zívání slyšet, ale není vidět (Arnott a kol., 2009; Massen a kol., 2015).
U lidí je jejich fylogeneticky nejbližší druh opice (šimpanz: Pan paniscus ; bonobos: Pan troglodytes) a africká opice Theropithecus gelada, nakažlivé zívání je nejen přítomné (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; ale viz: Amici et al., 2014), ale také sociálně modulovaný, protože reakce na zívání je nejvyšší, pokud jsou zapojeny určité kategorie jednotlivců (např. příbuzní, členové skupiny, dominantní osoby; Palagi et al., 2009; Campbell a de Waal, 2011, 2014; Norscia a Palagi, 2011; Demuru a Palagi, 2012; Massen et al., 2012). Byly předloženy dva hlavní argumenty vysvětlující tuto sociální asymetrii v nakažlivém zívání, které byly seskupeny do dvou hlavních hypotéz: hypotéza emočního zkreslení (EBH), spojující nakažlivé zívání s emočním přenosem a hypotéza pozorování zaujatosti (ABH), která zohledňuje nakažlivé zívání jako motorická odezva, která podléhá rozdílům v procesech pozornosti shora dolů (Palagi et al., 2020).
EBH předpovídá, že sociální asymetrie pozorovaná v mírách zející nákazy odráží rozdíly v různých sociální vazby, zástupce emocionálních vazeb mezi jednotlivci. Tato hypotéza je podpořena důkazy, že míra nákazy zívnutí sleduje empatický trend (sensu Preston a de Waal, 2002), který je nejvyšší mezi jednotlivci sdílejícími nejsilnější emoční pouto. Konkrétně Norscia a Palagi (2011) zjistili, že u lidí je míra nákazy zívání největší v reakci na příbuzné a přátele než v reakci na známé a cizí lidi. U dospělých šimpanzů je nákaza zívnutí vyšší mezi skupinami ve srovnání s členy mimo skupinu (Campbell a de Waal, 2011) a u bonobosů je míra nákazy zívání největší u jedinců, kteří se více sdružují (Demuru a Palagi, 2012). Ve srovnávacím šetření zahrnujícím jak lidi, tak bonobos, Palagi et al. (2014) zjistili, že míra nákazy zívání byla ovlivněna kvalitou vztahu mezi jednotlivci více než druhy, k nimž subjekty patřily. Navíc u lidí se zívání zvyšuje s věkem, kdy se schopnost identifikovat emoce ostatních zvyšuje a klesá se stářím, když tato schopnost klesá (Wiggers a van Lieshout, 1985; Anderson a Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen and Anderson, 2011; Bartholomew and Cirulli, 2014). Míra nákazy zívání roste od dětství do dospělosti také u šimpanzů (Madsen a Persson, 2013).
ABH předpovídá, že sociální asymetrie pozorovaná u nákazy zívnutí může být způsobena rozdíly v sociální, vizuální pozornosti (Massen a Gallup, 2017). Zejména nejvyšší úrovně nakažlivého zívání by byly způsobeny extra selektivní vizuální pozorností věnovanou jednotlivcům, kteří jsou pro pozorovatele relevantnější, například známým subjektům, jak se to vyskytuje u lidí a gelad, nebo dominancí, protože vyskytuje se u šimpanzů nebo bonobů (Yoon a Tennie, 2010; Massen et al., 2012; Massen and Gallup, 2017).Podle Massena a Gallupa (2017) by ABH byla podpořena existujícími důkazy o odlišné vizuální detekci a vizuálním vnímavém kódování tváří známých nebo skupinových subjektů ve srovnání s neznámými (např. Buttle a Raymond, 2003 ; Ganel a Goshen-Gottstein, 2004; Jackson a Raymond, 2006; Michel et al., 2006).
V této studii jsme analyzovali údaje o zívnutí shromážděné po dobu 9 let na lidech v jejich přirozeném prostředí a extrapolovali jsme případy, kdy zívnutí emitované subjektem bylo slyšet, ale neviděl potenciální respondent (sluchové zívání). Zvažováním pouze případů, kdy nebylo možné zjistit vizuální podnět zívajícího podnětu, jsme ověřili, zda sociální asymetrie dříve pozorovaná u míry zívnutí přetrvávala nebo ne. Testovali jsme zejména následující alternativní předpovědi odvozené ze dvou výše uvedených hypotéz (EBH a ABH).
Predikce 1a: podle EBH jsou míry nákazy zívnutí ovlivněny silou inter -individuální sociální vazba – zastoupení emocionální vazby – sama o sobě, a nikoli jinou, shora vybranou, selektivní vizuální pozorností věnovanou konkrétním jednotlivcům. Je-li tato hypotéza podporována, očekáváme, že budeme pozorovat sociální zkreslení, i když je vyloučena vizuální narážka na zívající podnět a míra sluchového nakažlivého zívání bude vyšší mezi silně vázanými ve srovnání se slabě vázanými jednotlivci.
Predikce 1b: podle ABH by vyšší úrovně nákazy zívání mezi silně vázanými jedinci ve srovnání se slabě vázanými jedinci byly spojeny s nejbližší selektivní vizuální pozorností shora dolů, kterou jednotlivci věnují jednotlivcům, kteří jsou pro ně relevantní, např. Rodina a přátelé. Pokud je tato hypotéza podporována, sociální zkreslení pozorované v nákaze zívnutí by mělo zmizet, když se uvažuje pouze o zívnutí sluchu, protože vizuální narážka se potenciálního respondenta nemůže zúčastnit.
Materiály a metody
Sběr dat a provozní definice
V této studii jsme uvažovali o vokalizovaných zívnutích vydávaných subjektem, které mohl slyšet – ale neviděl – pouze potenciální respondent (dále jen: sluchové zívnutí). Vysílač a potenciální respondenti museli být v rozmezí ≤5 m. Vokalizované zívání zahrnovalo použití hlasivek a zívání, která zahrnovala pouze těžkou inspiraci / expiraci, nebyly považovány za vokalizované; 294 případů sluchových zívnutí bylo extrapolováno z datového souboru celkem 2001 záchvatů zívání shromážděných během 9 let – od roku 2010 do roku 2019 – pomocí metody vzorkování všech výskytů (Altmann, 1974). Konkrétně sluchové zívnutí se shromažďovalo od listopadu 2010 do května 2019, od 05:30 do 02:30, na lidských bělošských subjektech ve věku od 18 do 77 let během jejich rutinních činností, např. Na pracovištích, při jídle, při společenských setkáních atd. , přičemž subjekty nevěděly, že budou pozorovány, a bez jakéhokoli zjevného vnějšího zdroje úzkosti. Databáze sluchového zívnutí obsahovala 193 dyadů reagujících na zívnutí. V závislosti na situaci byly informace zaznamenány, bez povšimnutí, pomocí alfanumerických kódů a vloženy přímo do výpočtových listů, zadány do mobilních telefonů nebo napsány na papír a poté vloženy do výpočtových listů pro následné zpracování. Autoři věděli o základních informacích, jako je věk a vztah mezi lidmi. Potenciální respondenti byli kódováni jako ve stavu neviditelnosti, když byla jejich hlava otočena o 180 ° vzhledem ke spoušti nebo když byla přítomna fyzická překážka blokující zrak, která potenciálnímu respondentovi bránila v pohledu na tvář a tělo spouště. Spouštěč a respondér nebyli nikdy úplně izolováni (např. Ve dvou samostatných místnostech se zavřenými dveřmi) od sebe navzájem. Sociální blízkost byla shromážděna na čtyřech úrovních: 0 = cizinci, kteří se nikdy předtím nesetkali; 1 = známí, kteří sdíleli výhradně nepřímý vztah založený na třetím vnějším prvku, tj. Pracovní povinnost (kolegové) nebo společní přátelé (přátelé přátel); 2 = přátelé, nesouvisející jednotlivci sdílející přímý vztah, který se netýká výlučně třetího vnějšího prvku; 3 = stálí partneři a příbuzní (r ≥ 0,25). Předchozí literatura uvádí, že odpovědi na zívnutí lze vyvolat do 5 minut po sledování zívnutí někoho jiného (spouštěcí zívnutí) (Provine, 1986), s maximem v první minutě (Provine, 2005; Palagi et al., 2014). Literatura také uvádí, že od čtvrté minuty existuje nejvyšší pravděpodobnost autokorelace (což znamená, že přítomnost zívnutí prováděného subjektem v čase t0 zvyšuje pravděpodobnost dalšího zívnutí stejným subjektem v čase t (0 + X), kde X je rostoucí jednotka času; Kapitány a Nielsen, 2017). Zvažovali jsme tedy odpovědi na zívnutí, které se vyskytly v časovém okně 3 minuty od zívnutí emitovaného spouštěčem.Pro další snížení zkreslení autokorelace jsme v případě řetězce zívnutí emitovaných spouštěčem (více zívnutí emitovaných v časovém okně 3 minuty) zaregistrovali jako odpověď pouze první zívnutí provedené po vnímání posledního zívnutí. Kódovali jsme zívnutí jako „spontánní“, když za 5 minut před událostí zívnutí nezíval žádný jiný subjekt.
Statistické analýzy
U analýz byly brány v úvahu následující proměnné: výskyt nákazy, kódováno jako: 1 = přítomnost, 0 = absence; sociální pouto bylo zadáno ve čtyřech výše definovaných úrovních (0 = cizinci; 1 = známí; 2 = přátelé; 3 = příbuzní); pohlaví spouštěče a pozorovatele bylo označeno jako : M = muž, F = žena; věkové třídy spouštěče a respondenta byly kódovány následovně: yo = mládež (18–24 let); ad = dospělý (25–64 let); se = senior (výše 65 let) (Statistics Canada, 2009); časové úseky byly kódovány takto: 1 = 05: 30–09: 00; 2 = 09:01–12: 30; 3 = 12: 31–16: 00:00; 4 = 16: 01–19: 30; 5 = 19: 31–23: 00; 6 = 23: 01–02: 30 (Giganti a Zilli, 2011). Databáze (viz doplňkový datový list) zahrnovalo 84 mužů, 69 žen, 16 mladých lidí (yo), 122 dospělých (ad) a 15 seniorů (se). Vyzkoušet, zda je výskyt zívnutí nákaza byla ovlivněna vazbou faktorů (0 = cizinci; 1 = známí; 2 = přátelé; 3 = příbuzní), pohlaví spouštěče (Trigger_sex), pohlaví respondenta (Responder_sex), věková třída spouštěče (Trigger_ageclass) a věková třída respondenta (Responder_ageclass) a časový úsek (od 1 do 6) ), jsme použili zobecněný lineární smíšený model (GLMM), který zahrnoval těchto pět prediktorů jako fixní efekty a spouštěče (Trigger) a respondenty (Responder) identity jako náhodné efekty. Modely jsme osadili do R (R Core Team, 2018; verze 3.5.1) pomocí funkce lmer balíčku R lme4 (Bates et al., 2015). Zjistili jsme význam celého modelu porovnáním s nulovým modelem zahrnujícím pouze náhodné efekty (Forstmeier a Schielzeth, 2011). K testování této významnosti jsme použili test poměru pravděpodobnosti (Dobson, 2002) (ANOVA s argumentem „Chisq“). Hodnoty p pro jednotlivé prediktory jsme vypočítali na základě testů poměru pravděpodobnosti mezi úplným a nulovým modelem pomocí R -funkce „drop1“ (Barr et al., 2013). Protože proměnná odezvy byla binární, použili jsme binomické rozdělení chyb. Testovali jsme, zda interakce mezi pohlavími nebo věkovými třídami spouštěče a respondenta byla významná, ale protože tomu tak nebylo, nezahrnuli jsme je do modelu. Použili jsme balíček vícenásobného kontrastu (multcomp) k provedení všech párových srovnání pro každou úroveň vazby pomocí testu Tukey (Bretz et al., 2010). Uváděli jsme Bonferroni upravené hodnoty p, odhad (Est), standardní chybu (SE) a hodnoty z.
Výsledky

Obrázek 1. Čárový graf vlivu sociální vazby mezi spouštěčem a respondérem (osa X) na průměrný výskyt nákazy akustickým zívnutím (osa Y). Přátelé a příbuzní vykazují významně vyšší frekvence nákazy zívnutí než cizí a známí (Tukeyův test: přátelé versus cizí lidé p < 0,001; příbuzní proti cizím lidem p < 0,001; přátelé versus známí p < 0,001; příbuzní versus známí p < 0,001; jiné kombinace, ns).
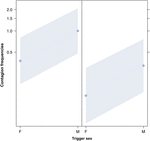
Obrázek 2. Čárový graf účinek pohlaví spouštěče (osa X) na průměrný výskyt nákazy zíváním (osa Y). Vliv spouštěcího pohlaví na nákazu zíváním, když respondérem je žena (vpravo) a muž (vlevo). Zívání mužů vyvolává více zívnutí než žen, bez ohledu na pohlaví respondenta (výsledek GLMM, p = 0,022). Body představují účinek, který má proměnná odezvy na nezávislou proměnnou „pohlaví spouštěče“, na základě hodnoty předpovězené modelem. Barevné pásy ukazují 95% interval spolehlivosti.
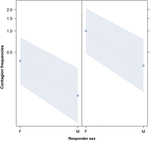
Obrázek 3. Čárový graf účinku pohlaví respondér (osa X) na průměrný výskyt nákazy zíváním (osa Y). Vliv pohlaví respondenta na nákazu akustického zívnutí, když je spouštěč žena (vpravo) nebo muž (vlevo). Ženy reagují výrazně více než muži bez ohledu na to pohlaví spouštěče (výsledek GLMM, p = 0,021). Body představují účinek, který má proměnná odezvy na nezávislou proměnnou „pohlaví respondenta“, na základě hodnoty předpovězené modelem. Barevné pruhy ukazují 95% interval spolehlivosti.
diskuse
Tato studie poprvé ukazuje, že nákaza zívání je významně ovlivněna sociální vazbou mezi jednotlivci (tabulka 1), i když spouštěcími podněty jsou sluchové zívání, které jsme definovali jako vokalizované zívání, které bylo možné slyšet, ale nevidět (vizuální podnět nezjistitelný, sluchový podnět detekovatelný). Zejména sluchové nakažlivé zívání je výrazně častější mezi příbuznými a přáteli než mezi cizími lidmi a známými (obrázek 1). Toto zjištění podporuje predikci 1a založenou na EBH, a nikoli predikci 1b založenou na ABH, což vede k závěru, že u lidí shora dolů nemůže být selektivní vizuální pozornost hlavní hnací silou sociální asymetrie pozorované u míry zívnutí (Norscia a Palagi , 2011; Norscia et al., 2016a). Rovněž pohlaví spouštěče a pohlaví přijímače mělo významný vliv na míru nákazy zívnutím, přičemž na muže – jako na spouštěče – reagovali častěji než ženy (obrázek 2) a ženy reagovaly častěji na zívnutí ostatních než muži (obrázek 3).
Na rozdíl od Bartoloměje a Cirulli (2014) jsme nezjistili žádný vliv věku na nákazu zívnutím, pravděpodobně proto, že naše databáze sluchových zívnutí měla silnou prevalenci dospělých (25–64 let) starý). Nejvyšší úrovně nákazy sluchového zívnutí u žen ve srovnání s muži potvrzují zkreslení pohlaví pozorované v naturalistických podmínkách u lidí náchylných k nákaze zívnutím Norscia et al. (2016a, b) při zvažování většího souboru dat, který zahrnoval také zívnutí, která by mohla být viděna potenciálním respondentem (se zíváním smyslové modality – vidění, sluchu nebo obojí – neovlivňující odpověď). Rovnost pohlaví je také v částečné shodě s předchozími výsledky získanými v kontrolovaných podmínkách, včetně vizuálních podnětů (Chan a Tseng, 2017; ale viz Norscia a Palagi, 2011; Bartholomew a Cirulli, 2014). Byla vyslovena hypotéza, že vysoký stupeň nákazy zíváním u žen může informovat o emoční nákaze (Norscia et al., 2016a), a to ve světle údajně vyšších empatických schopností – souvisejících s mateřstvím – žen ve srovnání s muži (Christov-Moore et al. ., 2014). O této otázce se však stále diskutuje, protože kulturní rozdíly mezi lidskými společnostmi mohou formovat dynamiku sociálních vazeb jiným způsobem. V této fázi poznání je proto komplikované oddělovat kulturní faktory, kvalitu mezilidských vztahů a vliv pohlaví při šíření nákazy zívnutím. Naše výsledky také ukazují, že muži fungují lépe než ženy jako spouštěče a nejšetrnější hypotézou pro to může být, že hlasové projevy mužů lze lépe slyšet v přirozeném prostředí, často charakterizovaném zvuky pozadí. Vnímání pohlaví hlasu se ve skutečnosti primárně opírá o základní frekvenci, která je v průměru nižší o oktávu v mužských než ženských hlasech, přičemž vokalizace s nižší frekvencí cestují dále než vysokofrekvenční (Marten a Marler, 1977; Latinus a Taylor, 2012). . Pokud je nám známo, neexistuje žádná konkrétní studie zabývající se možným zkreslením pohlaví ve slyšitelnosti zívnutí, a proto je pro ověření této spekulace nutné další zkoumání experimentálních studií v kontrolovaném stavu.
V této studii jsme také zjistili že rozdíly v míře nákazy zívnutí napříč kategoriemi (rodina a přátelé, cizí lidé a známí) nelze vysvětlit rozdíly v selektivní vizuální pozornosti shora dolů. Toto zjištění je v souladu s předchozí literaturou. Zdá se, že nakažlivé zívání zahrnuje oblasti mozku, které více souvisejí se sítí orientovaného zdola nahoru než s oblastmi souvisejícími shora dolů. Nákaza zívnutí navíc není ani citlivá na smyslové vjemy přítomné v signálu (sluchové, vizuální nebo audiovizuální) (Arnott et al., 2009; Norscia a Palagi, 2011), ani není ovlivněna vizuální perspektivou spouštěcího stimulu ( zívnutí v orientacích 90 °, 180 ° a 270 ° jsou schopna spouštět reakce zívání jako čelní zívání 0 °; Provine, 1989, 1996). Chan a Tseng (2017) zjistili, že schopnost detekovat zívnutí jako taková (percepční citlivost detekce) souvisela s délkou pohledu do očí obličeje uvolňujícího stimul, ale vzory očí nebyly schopny modulovat nakažlivé zívání. U šimpanzů byly nakažlivé frekvence zívání nejvyšší u jedinců ve stejné skupině než u jednotlivců z různých skupin, i když se respondenti dívali déle na videa mimo skupinu šimpanzů (Campbell a de Waal, 2011). Argument, že vizuální selektivní pozornost může ovlivnit míru nákazy zíváním v určitém směru (subjekty reagující spíše na rodinu než na cizince), je rovněž podkopáván absencí jakéhokoli konkrétního vzorce sociální pozornosti u člověka a primátů. Prostřednictvím sledování očí (aplikovaného na měření doby sledování) a zobrazováním neznámých tváří experimentálním subjektům Méary et al.(2014) zjistili, že lidé byli vychýlení k tváří vlastní rasy, zatímco pozornost makaků rhesus přitahovala více tváře nových než stejných druhů. Kawakami a kol. (2014) zjistili, že lidské subjekty věnovaly větší pozornost očím členů etnických skupin a nosu a ústům členů etnických skupin. Stejná studie také odhalila, že vizuální pozornost nezávisí na cílové rase. Měřením toho, jak dlouho experimentální subjekty hleděli na obrazovku, Whitehouse et al. (2016) poznamenali, že makakové Barbary věnovali větší pozornost škrábání videí cizinců než cizinců, ale také poznamenali, že v cizincích makakové věnovali největší pozornost těm jednotlivcům, se kterými sdíleli slabé sociální vazby. Schino a Sciarretta (2016) měřením rychlosti pohledů zjistili, že mandrily se zaměřovaly více na své vlastní příbuzné než na jiné než příbuzné, ale také více na dominantní než na podřízené skupinové kamarády. Proto tyto studie (používané k podpoře ABH) nepopisují žádný jediný vzor selektivní pozornosti. Jedním dalším důležitým bodem, který je třeba vzít v úvahu, je samotná definice známosti a členství ve skupině přijatá většinou studií používaných na podporu ABH (Massen and Gallup, 2017). Tyto studie ukázaly lepší vizuální detekci a vizuální vnímání kódování tváří známých / skupinových subjektů ve srovnání s neznámými, ale definovaly známost a členství ve skupině ne na základě osobních vztahů mezi jednotlivci. Místo toho byla známost nebo členství ve skupině definováno na základě nepřímých znalostí (např. Fotografie slavných lidí nebo subjektu již ukázaného ve fázi před zahájením soudního řízení) nebo společné rasy (např. Buttle a Raymond, 2003; Ganel a Goshen- Gottstein, 2004; Jackson a Raymond, 2006; Michel et al., 2006). Tato definice je pro účely těchto studií v pořádku, ale není tak dobrá, pokud jsou výsledky použity k navržení alternativních vysvětlení vlivu, který může mít skutečné sociální pouto – založené na skutečných vztazích – na jev, v tomto případě zívání nákazy . Například Michel et al. (2006) zjistili, že bělošští a asijští jedinci mohou lépe rozpoznávat tváře stejných ras, ale tento rozdíl nebyl přítomen u asijských jedinců, kteří žili asi rok mezi bělochy. Další bod diskuse se týká přítomnosti nákazy zívnutí u dětí s poruchou autistického spektra (ASD), které často vykazují změny vizuální pozornosti (Richard a Lajiness-O’Neill, 2015). U dětí s ASD může nákaza zívnutí být nepřítomná (Senju et al., 2007), narušena (Helt et al., 2010) nebo podobná typicky se vyvíjejícím dětem, když jsou subjekty indukovány k přesměrování své pozornosti video stimul během experimentálních pokusů (Usui et al., 2013). V nedávné studii Mariscal et al. (2019) zjistili, že nákaza zívání u dětí s ASD pozitivně souvisí s koncentrací oxytocinu v krvi, což je hormon zapojený do rodičovské a sociální vazby (Decety et al., 2016), a předpokládá, že nákaza zívání u dětí s ASD může souviset s variabilním průměrem koncentrace oxytocinu v různých studijních kohortách (Mariscal et al., 2019). Toto zjištění je v souladu s hypotézou EBH, která spojuje míru nákazy zívnutí se sociálními vazbami, které mohou odrážet emoční vazby.
Závěr
Naše studie přispívá k diskusi o mechanismech, které jsou základem sociální asymetrie v nákaze zívnutí (kritické recenze: viz Adriaense et al., 2020; Palagi et al., 2020) tím, že ukazuje, že nákaza zívání je pravděpodobně spojena spíše se selektivní pozorností zdola nahoru než se shora dolů. Pozornost zdola nahoru je primárně vedena smyslovým vnímáním vyvolávajícího podnětu, zatímco selektivní pozornost shora dolů je dobrovolný, trvalý proces, při kterém je interně vybrána konkrétní položka, která je zaměřena nebo zkoumána (Katsuki a Constantinidis, 2014). V tomto ohledu byl slyšet akustický stimul (sluchové zívnutí) vydávaný spouštěčem, který mohl vyvolat zívající odpověď v přijímači, i když přijímač nevěnoval spouštěči žádnou dobrovolnou vizuální pozornost. Míra odpovědi na zívání byla navíc sociálně modulována, přičemž nákaza zíváním byla nejvyšší u jedinců, kteří byli k sobě navzájem nejsilněji vázáni. Selektivní pozornost shora dolů tedy není hlavní hnací silou sociální asymetrie pozorované u nákazy zívnutí, což se jeví jako fenomén stimulovaný v souvislosti s procesy pozornosti zdola nahoru. Je zapotřebí dalšího zkoumání, abychom pochopili, zda a jakým způsobem mohou jiné formy pozornosti nebo předběžné fáze ovlivnit zívání.
Prohlášení o dostupnosti dat
Soubor dat použitý pro tuto studii je uveden v doplňkovém materiálu.
Etické prohlášení
Studie zahrnující lidské účastníky byla přezkoumána a schválena Comitato di Bioetica dAteneo (University Bioethical Committee) – University of Turin (ref č. 451945).V souladu s vnitrostátními právními předpisy a institucionálními požadavky nebyl pro tuto studii vyžadován písemný informovaný souhlas s účastí.
Příspěvky autorů
IN a EP provedly sběr dat a vytvořily a napsaly rukopis. AZ pomohla se sběrem dat a revizí rukopisů, čísly a tabulkami. Společnost MG provedla statistické analýzy a napsala související část rukopisu.
Konflikt zájmů
Autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly lze chápat jako potenciální střet zájmů.
Poděkování
Autoři by rádi poděkovali profesorům Roberto Barbuti a Prof. Cristina Giacoma za podporu výzkumu na univerzitě v Pise a Univerzita v Turíně.
Doplňkový materiál
Altmann, J. (1974). Pozorovací studie chování: metody odběru vzorků. Chování 49, 227–266. doi: 10.1163 / 156853974×00534
PubMed Abstract | CrossRef Full Text | Google Scholar
Anderson, J. R. a Meno, P. (2003). Psychologické vlivy na zívání u dětí. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J. a Cirulli, E. T. (2014). Individuální variace na nakažlivou náchylnost k zívání jsou vysoce stabilní a do značné míry nevysvětlitelné empatií nebo jinými známými faktory. PLoS One 9: e91773. doi: 10,1371 / journal.pone.0091773
PubMed Abstract | CrossRef Full Text | Google Scholar
Bretz, F., Hothorn, T. a Westfall, P. (2010). Několik srovnání pomocí R. Boca Ratona, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Etologická studie zívání primátů. I. Kvantitativní analýza a studium příčinných souvislostí u dvou druhů opic Starého světa (Cercocebus albigena a Macaca fascicularis). Ethology 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Full Text | Google Scholar
Dobson, A. J. (2002). An Introduction to Generalized Linear Models, 2nd Edn. Boca Raton, FL: Chapman and Hall.
Google Scholar
Katsuki, F. a Constantinidis, C . (2014). Pozornost zdola nahoru a shora dolů: různé procesy a překrývající se neurální systémy. Neurovědec 20, 509–521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef Full Text | Google Scholar
Singer, T. (2006). Neuronální základ a ontogeneze empatie a čtení mysli: přehled literatury a důsledky pro budoucí výzkum. Neurosci. Biobehav. Rev. 30, 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Statistics Canada (2009). Věkové kategorie, seskupení životního cyklu. Ottawa: Statistics Canada.
Google Scholar