Wprowadzenie
Ziewanie to mimowolna sekwencja otwierania ust, głębokiego wdechu, krótkiego bezdechu i mniej lub bardziej powolnego wydechu (Baenninger, 1997; Walusinski i Deputte, 2004; Guggisberg i in., 2010; Krestel i in., 2018). Po wywołaniu ziewnięcia nie można całkowicie stłumić. Dlatego został zdefiniowany jako wzorzec stereotypowy lub odruchowy (Lehmann, 1979; Provine, 1986). W Homo sapiens postawiono kilka hipotez ze zmiennym wsparciem wyjaśniającym mechanizmy i funkcje spontanicznego ziewania, takie jak utlenowanie (funkcja oddechowa spowodowana niedotlenieniem), zachowania związane ze stresem (wywołane pobudzeniem) lub termoregulacja (spowodowana hipertermią; Guggisberg i in., 2010; Massen i in., 2014; Gallup i Gallup, 2019). Będąc reakcją fizjologiczną, na ziewanie mogą wpływać czynniki wewnętrzne i zewnętrzne, takie jak pora dnia (Giganti i Zilli, 2011) lub temperatura wewnątrzczaszkowa / mózgowa (Gallup i Eldakar, 2013).
Ziewanie może kierować się na siebie i / lub pokazywać innym (Moyaho i in., 2017; Palagi i in., 2019). U ludzi i innych naczelnych, w zależności od gatunku, pokazywanie ziewania innym może powodować zagrożenie (Troisi i in., 1990; Deputte, 1994) i / lub zmiany fizjologiczne i behawioralne (Provine i in., 1987 ; Leone i in., 2015; Zannella i in., 2015). U ludzi ziewanie jest reakcją modulowaną społecznie, ponieważ może być hamowane przez rzeczywistą – a nie wirtualną – obecność społeczną (Gallup i in., 2019) oraz ponieważ ziewanie może być wywołane przez ziewanie kogoś innego w wyniku zjawiska znanego jako zaraźliwe ziewanie (Provine, 1989, 2005). Zakażenie ziewaniem można wywołać, nawet jeśli ziewanie jest słyszalne, ale niewidoczne (Arnott i in., 2009; Massen i in., 2015).
U ludzi ich filogenetycznie najbliższe gatunki małp (szympansy: Pan paniscus ; bonobos: Pan troglodytes) i afrykańskiej małpy Theropithecus gelada, ziewanie zakaźne jest obecne nie tylko (Provine, 1986; Palagi i in., 2009; Tan i in., 2017; ale patrz: Amici i in., 2014), ale także modulowane społecznie, ponieważ reakcja ziewania jest najwyższa, gdy zaangażowane są pewne kategorie osób (np. krewniacy, członkowie grupy, dominanty; Palagi i in., 2009; Campbell i de Waal, 2011, 2014; Norscia i Palagi, 2011; Demuru i Palagi, 2012; Massen i in., 2012). Przedstawiono dwa główne argumenty wyjaśniające tę społeczną asymetrię w zaraźliwym ziewaniu, które zgrupowano w dwie główne hipotezy: hipotezę uprzedzenia emocjonalnego (EBH), łączącą zaraźliwe ziewanie z transferem emocjonalnym oraz hipotezę uprzedzeń uważności (ABH), która uwzględnia zaraźliwe ziewanie jako reakcja motoryczna, która podlega różnicom w odgórnych procesach uwagi (Palagi i in., 2020).
EBH przewiduje, że asymetria społeczna obserwowana we współczynnikach zarażenia ziewaniem odzwierciedla różnice w różnych więzi społeczne, zastępstwo więzi emocjonalnej między jednostkami. Hipoteza ta jest poparta dowodami, że wskaźniki zarażenia ziewaniem podążają za tendencją empatyczną (sensu Preston i de Waal, 2002), która jest najwyższa między osobami łączącymi najsilniejszą więź emocjonalną. W szczególności Norscia i Palagi (2011) stwierdzili, że u ludzi wskaźniki zarażenia ziewaniem są największe w odpowiedzi na krewnych i przyjaciół niż w odpowiedzi na znajomych i nieznajomych. U dorosłych szympansów zarażenie ziewaniem jest wyższe między członkami grupy w porównaniu z członkami z grup zewnętrznych (Campbell i de Waal, 2011), a u bonobo wskaźniki zarażenia ziewaniem są największe między osobnikami, które są ze sobą bardziej powiązane (Demuru i Palagi, 2012). W badaniu porównawczym obejmującym zarówno ludzi, jak i bonobo, Palagi i wsp. (2014) stwierdzili, że na wskaźniki zarażenia ziewaniem w większym stopniu wpływa jakość relacji między osobnikami niż gatunek, do którego należeli badani. Dodatkowo u ludzi zarażenie ziewaniem wzrasta wraz z wiekiem, gdy zdolność rozpoznawania emocji innych wzrasta i maleje wraz ze starością, gdy taka zdolność spada (Wiggers i van Lieshout, 1985; Anderson i Meno, 2003; Saxe i in., 2004; Singer, 2006; Millen i Anderson, 2011; Bartholomew i Cirulli, 2014). Wskaźniki zarażenia ziewaniem rosną od niemowlęctwa do dorosłości także u szympansów (Madsen i Persson, 2013).
ABH przewiduje, że asymetria społeczna obserwowana w zarażaniu ziewaniem może wynikać z różnic w społecznej i wzrokowej uwadze (Massen i Gallup, 2017). W szczególności najwyższe poziomy zaraźliwego ziewania wynikałyby z dodatkowej odgórnej, selektywnej uwagi wzrokowej zwracanej na osoby, które są bardziej istotne dla obserwatora, takie jak znane przedmioty, jak to występuje u ludzi i gelad, lub dominujących, ponieważ występuje u szympansów lub bonobo (Yoon i Tennie, 2010; Massen i in., 2012; Massen i Gallup, 2017).Według Massena i Gallupa (2017), ABH byłby poparty istniejącymi dowodami na temat innego wykrywania wizualnego i wizualnego kodowania percepcji twarzy osób znajomych i / lub osób z grupy w porównaniu z nieznanymi (np. Buttle i Raymond, 2003 ; Ganel i Goshen-Gottstein, 2004; Jackson i Raymond, 2006; Michel i in., 2006).
W tym badaniu przeanalizowaliśmy dane dotyczące ziewania zebrane w ciągu 9 lat na temat ludzi w ich naturalnym środowisku i ekstrapolowaliśmy przypadki, w których ziewanie emitowane przez podmiot było słyszalne, ale nie widziane przez potencjalnego ratownika (ziewanie słuchowe). Rozważając tylko te przypadki, w których wizualny sygnał bodźca ziewania nie był wykrywalny, zweryfikowaliśmy, czy asymetria społeczna obserwowana wcześniej we wskaźnikach zarażenia ziewaniem utrzymywała się, czy nie. W szczególności przetestowaliśmy następujące alternatywne prognozy wyprowadzone z dwóch przedstawionych powyżej hipotez (EBH i ABH).
Prognoza 1a: zgodnie z EBH, na stopień zarażenia ziewania ma wpływ siła inter -indywidualna więź społeczna – proxy więzi emocjonalnej – per se, a nie przez inną odgórną, selektywną uwagę wzrokową skierowaną w szczególności na określone osoby. Jeśli ta hipoteza zostanie potwierdzona, spodziewamy się zaobserwować stronniczość społeczną również wtedy, gdy wyklucza się wizualną wskazówkę bodźca ziewania, a wskaźniki zaraźliwego ziewania słuchowego będą wyższe między osobami silnie związanymi w porównaniu do osób słabo związanych.
Prognoza 1b: zgodnie z ABH wyższy poziom zarażenia ziewaniem między silnie związanymi w porównaniu z osobami słabo związanymi byłby powiązany z najbliższą odgórną, selektywną uwagą wzrokową, jaką osoby zwracają na osoby, które są dla nich istotne, np. Rodzina i przyjaciele. Jeśli ta hipoteza jest potwierdzona, społeczne uprzedzenia obserwowane w przypadku zarażenia ziewaniem powinny zniknąć, gdy bierze się pod uwagę tylko ziewnięcia słuchowe, ponieważ potencjalny ratownik nie może uczestniczyć w sygnale wizualnym.
Materiały i metody
Zbieranie danych i definicje operacyjne
W tym badaniu wzięliśmy pod uwagę głośne ziewnięcia emitowane przez podmiot, który mógł być słyszany – ale niewidoczny – przez potencjalnego ratownika (dalej: ziewanie słuchowe). Emiter i potencjalni reagujący musieli znajdować się w odległości ≤5 m. Wokalizowane ziewnięcia wymagały użycia fałdów głosowych, a ziewnięcia, które obejmowały tylko ciężki wdech / wydech, nie były uważane za wokalizowane; 294 przypadki ziewania słuchowego ekstrapolowano ze zbioru danych obejmującego ogółem ataków ziewania z 2001 roku zebranych w ciągu 9 lat – od 2010 do 2019 roku – przy użyciu metody próbkowania wszystkich zdarzeń (Altmann, 1974). W szczególności ziewnięcia słuchowe zbierano od listopada 2010 r. Do maja 2019 r., Od 05:30 do 02:30, u osób rasy kaukaskiej w wieku od 18 do 77 lat podczas ich rutynowych czynności, np. W miejscach pracy, podczas posiłków, podczas spotkań towarzyskich itp. ., przy czym badani byli nieświadomi bycia obserwowanymi i przy braku jakiegokolwiek widocznego zewnętrznego źródła niepokoju. Baza danych ziewania słuchowego zawierała 193 diady reagujących na potencjalne ziewanie. W zależności od sytuacji informacje były zapisywane bez adnotacji za pomocą kodów alfanumerycznych i wpisywane bezpośrednio do arkuszy kalkulacyjnych, wpisywane w telefonach komórkowych lub zapisywane na papierze, a następnie wprowadzane do arkuszy kalkulacyjnych w celu późniejszego opracowania. Podstawowe informacje, takie jak wiek i relacje międzyludzkie, były znane autorom. Potencjalni ratownicy zostali zakodowani tak, jak w stanie niewidzącym, kiedy ich głowa została obrócona o 180 ° w stosunku do spustu lub gdy obecna była fizyczna przeszkoda blokująca wzrok, uniemożliwiająca potencjalnemu ratownikowi zobaczenie twarzy i ciała wyzwalacza. Wyzwalacz i osoba odpowiadająca nigdy nie były całkowicie odizolowane (np. W dwóch oddzielnych pokojach z zamkniętymi drzwiami) od siebie. Bliskość społeczną zebrano na czterech poziomach: 0 = obcy, którzy nigdy wcześniej się nie spotkali; 1 = znajomi, których łączyła wyłącznie relacja pośrednia oparta na trzecim elemencie zewnętrznym, czyli obowiązkach zawodowych (koledzy) lub wspólnych znajomych (przyjaciele znajomych); 2 = przyjaciele, niespokrewnione osoby mające bezpośredni związek nie związany wyłącznie z trzecim zewnętrznym elementem; 3 = stali partnerzy i krewni (r ≥ 0,25). Wcześniejsze publikacje donosiły, że reakcje ziewania można wywołać w ciągu 5 minut po obejrzeniu czyjegoś ziewnięcia (ziewnięcie wyzwalacza) (Provine, 1986), z maksimum w pierwszej minucie (Provine, 2005; Palagi i in., 2014). W literaturze podaje się również, że od czwartej minuty istnieje najwyższe prawdopodobieństwo autokorelacji (co oznacza, że obecność ziewania wykonanego przez badanego w czasie t0 zwiększa prawdopodobieństwo ponownego ziewnięcia tego samego badanego w czasie t (0 + X), gdzie X jest rosnąca jednostka czasu; Kapitány i Nielsen, 2017). Dlatego rozważaliśmy reakcje na ziewanie występujące w ciągu 3 minut od ziewnięcia emitowanego przez wyzwalacz.Aby jeszcze bardziej zmniejszyć odchylenie autokorelacji, w przypadku łańcucha ziewnięć emitowanego przez wyzwalacz (więcej ziewów emitowanych w 3-minutowym oknie czasowym) zarejestrowaliśmy jako odpowiedź tylko pierwsze ziewanie wykonane po odczuciu ostatniego ziewnięcia. Zakodowaliśmy ziewanie jako „spontaniczne”, gdy żaden inny podmiot nie ziewnął w ciągu 5 minut poprzedzających ziewanie.
Analizy statystyczne
W analizach wzięto pod uwagę następujące zmienne: występowanie zarażenia, zakodowane jako: 1 = obecność, 0 = nieobecność; więź społeczna została wprowadzona na czterech poziomach zdefiniowanych powyżej (0 = obcy; 1 = znajomi; 2 = przyjaciele; 3 = krewniacy); płeć wyzwalacza i obserwatora została oznaczona jako : M = mężczyzna, F = kobieta; klasy wiekowe wyzwalacza i respondenta zakodowano następująco: yo = młodzież (18–24 lata); ad = dorosły (25–64 lata); se = senior (powyżej 65 lat) (Statistics Canada, 2009); przedziały czasowe zostały zakodowane w następujący sposób: 1 = 05: 30–09: 00; 2 = 09:01–12: 30; 3 = 12: 31–16: 00 po południu; 4 = 16: 01–19: 30; 5 = 19: 31–23: 00; 6 = 23: 01–02: 30 (Giganti i Zilli, 2011). Baza danych (patrz Dodatkowa karta danych) obejmowało 84 mężczyzn, 69 kobiet, 16 młodych ludzi (lat), 122 osoby dorosłe (reklama) i 15 osób starszych (osoby dorosłe). Aby sprawdzić, czy na zarażenie miały wpływ czynniki więzi (0 = obcy; 1 = znajomi; 2 = przyjaciele; 3 = krewniacy), płeć wyzwalacza (Trigger_sex), płeć osoby odpowiadającej (Responder_sex), klasa wiekowa wyzwalacza (Trigger_ageclass) i klasa wiekowa osoby odpowiadającej (Responder_ageclass) oraz przedział czasowy (od 1 do 6 ), użyliśmy uogólnionego liniowego modelu mieszanego (GLMM), który obejmował te pięć predyktorów jako efekty stałe i wyzwalacze (wyzwalacz) i tożsamości respondentów (osoba odpowiadająca) jako efekty losowe. Dopasowaliśmy modele w R (R Core Team, 2018; wersja 3.5.1) za pomocą funkcji lmer pakietu R lme4 (Bates i in., 2015). Istotność pełnego modelu ustaliliśmy przez porównanie z modelem zerowym zawierającym tylko efekty losowe (Forstmeier i Schielzeth, 2011). Użyliśmy testu współczynnika wiarygodności (Dobson, 2002), aby przetestować tę istotność (ANOVA z argumentem „Chisq”). Obliczono wartości p dla poszczególnych predyktorów w oparciu o testy współczynnika wiarygodności między modelem pełnym i zerowym przy użyciu metody R -funkcja „drop1” (Barr i in., 2013). Ponieważ zmienna odpowiedzi była binarna, użyliśmy dwumianowego rozkładu błędów. Sprawdziliśmy, czy interakcje między płciami lub klasami wieku wyzwalacza i odpowiadającego były istotne, ale ponieważ nie były, nie uwzględniliśmy ich w modelu. Użyliśmy pakietu wielu kontrastów (multcomp), aby wykonać wszystkie porównania parami dla każdego poziomu wiązania za pomocą testu Tukeya (Bretz i in., 2010). Podaliśmy wartości p, oszacowanie (Est), błąd standardowy (SE) i wartości z skorygowane przez Bonferroniego.
Wyniki

Rysunek 1. Wykres liniowy wpływu więzi społecznej między wyzwalaczem a reagującym (oś X) na średnie występowanie zaraźliwości akustycznej ziewania (oś Y). Przyjaciele i krewni wykazują znacznie wyższą częstotliwość zarażania ziewaniem niż nieznajomi i znajomi (test Tukeya: przyjaciele kontra nieznajomi p < 0,001; krewni kontra nieznajomi p < 0,001; znajomi kontra znajomi p < 0,001; krewni kontra znajomi p < 0,001; inne kombinacje, ns).
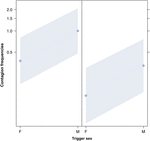
Rysunek 2. Wykres liniowy wpływ płci wyzwalacza (oś X) na średnie występowanie zaraźliwości akustycznej ziewania (oś Y). Wpływ płci wyzwalającej na zakaźne ziewanie akustyczne, gdy reagującym jest kobieta (po prawej) i mężczyzna (po lewej). Ziewnięcia mężczyzn wywołują więcej ziewnięć niż kobiety, niezależnie od płci osoby odpowiadającej (wynik GLMM, p = 0,022). Punkty reprezentują wpływ, jaki ma zmienna odpowiedzi na zmienną niezależną „płeć czynnika wyzwalającego” na podstawie wartości przewidywanej przez model. Kolorowe paski pokazują 95% przedział ufności.
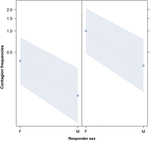
Rysunek 3. Wykres liniowy wpływu płci odpowiadający (oś X) na ziewanie akustyczne ziewanie średnie występowanie (oś Y). Wpływ płci osoby odpowiadającej na ziewanie akustyczne, gdy wyzwalaczem jest kobieta (po prawej) lub mężczyzna (po lewej). Kobiety reagują znacznie częściej niż mężczyźni, niezależnie od płci wyzwalacza (wynik GLMM, p = 0,021). Punkty reprezentują wpływ, jaki ma zmienna odpowiedzi na niezależną zmienną „płeć osoby odpowiadającej” na podstawie wartości przewidywanej przez model. Kolorowe prążki pokazują 95% przedział ufności.
Dyskusja
To badanie po raz pierwszy pokazuje, że na zarażenie ziewaniem istotny wpływ ma więź społeczna między osobami (Tabela 1), nawet jeśli bodźcem wyzwalającym są ziewnięcia słuchowe, które zdefiniowaliśmy jako ziewnięcia wokalne, które można było usłyszeć, ale nie można było ich zobaczyć (sygnał wzrokowy niewykrywalny, sygnał dźwiękowy wykrywalny). W szczególności słuchowe zaraźliwe ziewanie występuje znacznie częściej między krewnymi i przyjaciółmi niż między nieznajomymi i znajomymi (ryc. 1). Odkrycie to potwierdza prognozę 1a opartą na EBH, a nie prognozę 1b opartą na ABH, prowadząc do wniosku, że u ludzi odgórna selektywna uwaga wzrokowa nie może być głównym czynnikiem napędzającym asymetrię społeczną obserwowaną we wskaźnikach zarażenia ziewaniem (Norscia i Palagi , 2011; Norscia i in., 2016a). Również płeć wyzwalacza i odbierającego miały istotny wpływ na wskaźniki zarażenia ziewaniem, przy czym mężczyźni – jako wyzwalacze – reagowali częściej niż kobiety (ryc. 2), a kobiety częściej niż kobiety reagowały na ziewanie innych osób. mężczyzn (ryc. 3).
W przeciwieństwie do Bartholomew i Cirulli (2014), nie stwierdziliśmy wpływu wieku na zarażenie ziewaniem, najprawdopodobniej dlatego, że w naszej bazie danych o ziewaniach słuchowych występują osoby dorosłe (25–64 lat). stary). Najwyższe poziomy zakażenia słuchowego ziewaniem u kobiet w porównaniu z mężczyznami potwierdzają uprzedzenia płciowe obserwowane w warunkach naturalistycznych u ludzi podatnych na zarażenie ziewaniem przez Norscia i wsp. (2016a, b) biorąc pod uwagę większy zbiór danych, który zawierał także ziewnięcia, które może zobaczyć potencjalny reagujący (z ziewaniem modalności sensorycznej – wzrok, słuch lub jedno i drugie – bez wpływu na odpowiedź). Odchylenie ze względu na płeć jest również częściowo zgodne z poprzednimi wynikami uzyskanymi w kontrolowanych warunkach, w tym ze wskazówkami wizualnymi (Chan i Tseng, 2017; ale zobacz Norscia i Palagi, 2011; Bartholomew i Cirulli, 2014). Postawiono hipotezę, że wysoki stopień zarażenia ziewaniem u kobiet może wpływać na zarażenie emocjonalne (Norscia i in., 2016a), w świetle rzekomo wyższych zdolności empatycznych – związanych z macierzyństwem – kobiet w porównaniu z mężczyznami (Christov-Moore et al. ., 2014). Jednak ta kwestia jest nadal przedmiotem debaty, ponieważ różnice kulturowe w różnych społeczeństwach mogą w inny sposób kształtować dynamikę więzi społecznych. Dlatego na tym etapie wiedzy skomplikowane jest rozwikłanie czynników kulturowych, jakości relacji międzyludzkich i wpływu płci na rozprzestrzenianie się zakażenia ziewaniem. Nasze wyniki pokazują również, że mężczyźni działają lepiej niż kobiety jako czynniki wyzwalające, a najbardziej oszczędną hipotezą może być to, że wokalizacje mężczyzn są lepiej słyszalne w naturalnych warunkach, często charakteryzowanych przez hałas w tle. Rzeczywiście, percepcja płci głosu opiera się głównie na częstotliwości podstawowej, która jest średnio o oktawę niższa w głosach męskich niż żeńskich, z wokalizami o niższej częstotliwości przemieszczającymi się dalej niż wokalizacje o wysokiej częstotliwości (Marten i Marler, 1977; Latinus i Taylor, 2012) . Jednak, zgodnie z naszą wiedzą, nie ma konkretnego badania dotyczącego możliwego uprzedzenia ze względu na płeć w słyszalności ziewania i dlatego konieczne są dalsze badania z próbami eksperymentalnymi w kontrolowanych warunkach, aby zweryfikować te spekulacje.
W tym badaniu znaleźliśmy również że różnic we współczynniku zarażenia ziewaniem w różnych kategoriach (rodzina i przyjaciele, nieznajomi i znajomi) nie można wyjaśnić różnicami w odgórnej, selektywnej uwadze wzrokowej. To odkrycie jest zgodne z wcześniejszą literaturą. Wydaje się, że zaraźliwe ziewanie obejmuje obszary mózgu, które są bardziej związane z siecią zorientowaną, niż z obszarami odgórnymi. Co więcej, zarażenie ziewaniem nie jest wrażliwe na bodźce sensoryczne obecne w sygnale (słuchowe, wizualne lub audiowizualne) (Arnott i in., 2009; Norscia i Palagi, 2011) ani nie wpływa na wizualną perspektywę bodźca wyzwalającego ( ziewa w orientacji 90 °, 180 ° i 270 ° są w stanie wywołać reakcje ziewania, takie jak ziewanie czołowe, zerowe; Provine, 1989, 1996). Chan i Tseng (2017) stwierdzili, że zdolność do wykrywania ziewania jako takiego (czułość wykrywania percepcji) była związana z czasem patrzenia w oczy twarzy wyzwalającej bodziec, ale wzorce spojrzenia nie były w stanie modulować zaraźliwego ziewania. U szympansów częstość zaraźliwego ziewania była najwyższa między osobami z tej samej grupy niż osobnikami z różnych grup, nawet jeśli respondenci dłużej przyglądali się filmom szympansów z grup zewnętrznych (Campbell i de Waal, 2011). Argument, że selektywna uwaga wzrokowa może wpływać na ziewanie w określonym kierunku (pacjenci reagują bardziej na rodzinę niż na nieznajomych), jest również podważany przez brak określonego wzorca uwagi społecznej u ludzi i naczelnych innych niż ludzie. Poprzez śledzenie wzroku (stosowane do pomiaru czasu oglądania) i pokazywanie nieznanych twarzy badanym eksperymentalnym, Méary i in.(2014) zaobserwowali, że ludzie byli pochyleni w stronę twarzy własnej rasy, podczas gdy uwagę makaków rezusów bardziej przyciągały twarze nowe niż twarze tego samego gatunku. Kawakami i in. (2014) zaobserwowali, że ludzie zwracali większą uwagę na oczy członków grup etnicznych oraz nos i usta członków grup etnicznych. To samo badanie ujawniło również, że uwaga wzrokowa nie zależy od rasy docelowej. Mierząc, jak długo badani eksperymentalni patrzyli na ekran, Whitehouse i wsp. (2016) zaobserwowali, że makaki berberyjskie zwracały większą uwagę na drapanie filmów wideo z nieznajomymi niż osobami obcymi, ale zauważyli również, że wśród nieznajomych makaki zwracały największą uwagę na osoby, z którymi łączyły ich słabe więzi społeczne. Mierząc wskaźniki spojrzenia, Schino i Sciarretta (2016) zauważyli, że mandryle patrzyły bardziej na swoich krewnych niż na osoby niebędące krewnymi, ale także bardziej na dominujących niż na podwładnych kolegów z grupy. Dlatego te badania (wykorzystane na poparcie ABH) nie opisują żadnego pojedynczego wzorca selektywnej uwagi. Kolejną ważną kwestią do rozważenia jest sama definicja znajomości i przynależności do grupy przyjęta w większości badań wykorzystywanych do wspierania ABH (Massen i Gallup, 2017). Badania te wykazały lepsze wykrywanie wizualne i wizualne kodowanie percepcyjne twarzy osób znanych / w grupie w porównaniu z osobami nieznanymi, ale zdefiniowano zażyłość i przynależność do grupy nie na podstawie osobistych relacji między jednostkami. Zamiast tego, zażyłość lub przynależność do grupy definiowano na podstawie wiedzy pośredniej (np. Zdjęcie sławnych ludzi lub przedmiotu już pokazanego w fazie przedprocesowej) lub wspólnej rasy (np. Buttle i Raymond, 2003; Ganel i Goshen- Gottstein, 2004; Jackson i Raymond, 2006; Michel i in., 2006). Ta definicja jest w porządku dla celów tych badań, ale nie jest tak dobra, jeśli wyniki zostaną wykorzystane do zaproponowania alternatywnych wyjaśnień wpływu, jaki rzeczywiste więzi społeczne – oparte na prawdziwych związkach – mogą mieć na zjawisko, w tym przypadku zarażenie ziewaniem . Na przykład Michel i in. (2006) zaobserwowali, że osoby rasy białej i azjatyckiej mogły lepiej rozpoznawać twarze tej samej rasy, ale ta różnica nie była obecna u osób azjatyckich, które żyły przez około rok wśród rasy białej. Kolejny punkt dyskusji dotyczy obecności zarażenia ziewaniem u dzieci z zaburzeniami ze spektrum autyzmu (ASD), które często wykazują zmiany uwagi wzrokowej (Richard i Lajiness-O’Neill, 2015). U dzieci z ASD zarażenie ziewaniem może być nieobecne (Senju i in., 2007), upośledzone (Helt i in., 2010) lub podobne do typowo rozwijających się dzieci, gdy badani są nakłaniani do przekierowania uwagi na bodziec wideo podczas badań eksperymentalnych (Usui i in., 2013). W niedawnym badaniu Mariscal i wsp. (2019) stwierdzili, że zarażenie ziewaniem u dzieci z ASD było pozytywnie związane ze stężeniem oksytocyny we krwi, hormonu odpowiedzialnego za przywiązanie rodziców i społeczeństwa (Decety i in., 2016) i wysunęli wniosek, że zarażenie ziewaniem u dzieci z ASD może być związane ze zmienną średnią stężenia oksytocyny w różnych kohortach badawczych (Mariscal i wsp., 2019). To odkrycie jest zgodne z hipotezą EBH, która łączy współczynniki zarażenia ziewaniem z więziami społecznymi, które mogą odzwierciedlać więzi emocjonalne.
Wniosek
Nasze badanie uzupełnia dyskusję na temat mechanizmów leżących u podstaw asymetria społeczna w zarażaniu ziewaniem (krytyczne recenzje: patrz Adriaense i in., 2020; Palagi i in., 2020), pokazując, że zarażenie ziewaniem jest prawdopodobnie związane z oddolną, a nie odgórną, selektywną uwagą. Uwaga oddolna jest kierowana przede wszystkim przez zmysłową percepcję bodźca wywołującego, podczas gdy selektywna uwaga odgórna jest dobrowolnym, trwałym procesem, w którym dana pozycja jest wybierana wewnętrznie i skupiana na niej lub badana (Katsuki i Constantinidis, 2014). Pod tym względem bodziec akustyczny (ziewanie słuchowe) emitowany przez wyzwalacz był słyszalny i mógł wywołać w odbiorniku reakcję ziewania, nawet jeśli odbiornik nie zwracał świadomej uwagi wzrokowej na spust. Co więcej, wskaźniki reakcji ziewania były modulowane społecznie, przy czym zarażenie słuchowe było największe u osób, które były ze sobą najsilniej związane. Dlatego selektywna uwaga odgórna nie jest głównym motorem asymetrii społecznej obserwowanej w zarażeniu ziewaniem, które wydaje się być zjawiskiem stymulowanym przez bodźce związane z oddolnymi procesami uwagi. Konieczne są dalsze badania, aby zrozumieć, czy iw jaki sposób inne formy uwagi lub etapy pre-uważności mogą wpływać na zarażenie ziewaniem.
Oświadczenie o dostępności danych
Zbiór danych wykorzystany w tym badaniu znajduje się w materiale uzupełniającym.
Oświadczenie etyczne
Badanie z udziałem ludzi zostało przeanalizowane i zatwierdzone przez Comitato di Bioetica dAteneo (Uniwersytecka Komisja Bioetyczna) – Uniwersytet w Turynie (ref. . nr 451945).Pisemna świadoma zgoda na udział w tym badaniu nie była wymagana zgodnie z ustawodawstwem krajowym i wymogami instytucjonalnymi.
Wkład autorów
IN i EP zebrały dane, opracowały i napisały rękopis. AZ pomagał w zbieraniu danych i rewizji rękopisu, rycinach i tabelach. MG przeprowadził analizy statystyczne i napisał odpowiednią część pracy.
Konflikt interesów
Autorzy deklarują, że badanie zostało przeprowadzone przy braku jakichkolwiek powiązań handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Podziękowania
Autorzy pragną podziękować prof. Roberto Barbuti i prof. Cristinie Giacoma za wsparcie badań na Uniwersytecie w Pizie i Odpowiednio Uniwersytet w Turynie.
Materiał uzupełniający
Altmann, J. (1974). Obserwacyjne badanie zachowania: metody próbkowania. Behavior 49, 227–266. doi: 10.1163 / 156853974×00534
PubMed Abstract | Pełny tekst CrossRef | Google Scholar
Anderson, J. R., and Meno, P. (2003). Psychologiczne wpływy na ziewanie u dzieci. Curr. Psychol. Łotysz. 11, 1–7.
Google Scholar
Bartholomew, A. J. i Cirulli, E. T. (2014). Indywidualne zróżnicowanie podatności na zaraźliwe ziewanie jest wysoce stabilne i w dużej mierze niewyjaśnione przez empatię lub inne znane czynniki. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | Pełny tekst CrossRef | Google Scholar
Bretz, F., Hothorn, T. i Westfall, P. (2010). Wielokrotne porównania z udziałem R. Boca Ratona, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Badanie etologiczne ziewania u naczelnych. I. Analiza ilościowa i badanie związku przyczynowego u dwóch gatunków małp Starego Świata (Cercocebus albigena i Macaca fascicularis). Ethology 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Pełny tekst | Google Scholar
Dobson, A. J. (2002). Wprowadzenie do uogólnionych modeli liniowych, wyd. 2. Boca Raton, FL: Chapman and Hall.
Google Scholar
Katsuki, F. i Constantinidis, C . (2014). Uwaga oddolna i odgórna: różne procesy i nakładające się systemy neuronowe. Neuroscientist 20, 509–521. doi: 10.1177 / 1073858413514136
PubMed Abstract | Pełny tekst CrossRef | Google Scholar
Singer, T. (2006). Neuronalne podstawy i ontogeneza empatii i czytania w myślach: przegląd literatury i implikacje dla przyszłych badań. Neurosci. Biobehav. Rev. 30, 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | Pełny tekst CrossRef | Google Scholar
Statistics Canada (2009). Kategorie wiekowe, grupowanie cykli życia. Ottawa: Statistics Canada.
Google Scholar