Inleiding
Geeuwen is een onvrijwillige opeenvolging van mondopening, diepe inspiratie, korte apneu en min of meer langzame uitademing (Baenninger, 1997; Walusinski en Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Wanneer een geeuw wordt opgewekt, kan deze niet volledig worden onderdrukt. Daarom is het gedefinieerd als een stereotiep of reflexachtig patroon (Lehmann, 1979; Provine, 1986). In Homo sapiens zijn verschillende hypothesen naar voren gebracht met variabele ondersteuning om mechanismen en functies van spontaan gapen te verklaren, zoals oxygenatie (ademhalingsfunctie veroorzaakt door hypoxie), stressgerelateerd gedrag (veroorzaakt door opwinding) of thermoregulatie (veroorzaakt door hyperthermie; Guggisberg et al., 2010; Massen et al., 2014; Gallup en Gallup, 2019). Omdat het een fysiologische reactie is, kan gapen worden beïnvloed door interne en externe factoren, zoals het tijdstip van de dag (Giganti en Zilli, 2011) of intracraniële temperatuur / hersentemperatuur (Gallup en Eldakar, 2013).
Geeuwen kan wees zelfgestuurd en / of getoond aan anderen (Moyaho et al., 2017; Palagi et al., 2019). Bij menselijke en niet-menselijke primaten kan het, afhankelijk van de soort, wanneer anderen geeuwen, bedreiging (Troisi et al., 1990; Deputte, 1994) en / of fysiologische en gedragsveranderingen (Provine et al., 1987) communiceren. ; Leone et al., 2015; Zannella et al., 2015). Bij mensen is gapen een sociaal gemoduleerde reactie omdat het kan worden geremd door feitelijke – en niet virtuele – sociale aanwezigheid (Gallup et al., 2019) en omdat een geeuw kan worden veroorzaakt door de geeuw van iemand anders, als gevolg van een bekend fenomeen als besmettelijk gapen (Provine, 1989, 2005). Geeuwbesmetting kan zelfs worden uitgelokt als de geeuw wordt gehoord maar niet wordt gezien (Arnott et al., 2009; Massen et al., 2015).
Bij mensen is hun fylogenetisch meest nabije apensoort (chimpansees: Pan paniscus ; bonobos: Pan troglodytes) en de Afrikaanse aap Theropithecus gelada, is besmettelijk geeuwen niet alleen aanwezig (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; maar zie: Amici et al., 2014) maar ook sociaal gemoduleerd omdat de geeuwreactie het hoogst is wanneer bepaalde categorieën individuen erbij betrokken zijn (bijv. verwanten, groepsleden, dominanten; Palagi et al., 2009; Campbell en de Waal, 2011, 2014; Norscia en Palagi, 2011; Demuru en Palagi, 2012; Massen et al., 2012). Er zijn twee belangrijke argumenten aangevoerd om deze sociale asymmetrie bij besmettelijk gapen te verklaren, die zijn gegroepeerd in twee hoofdhypothesen: de Emotional Bias Hypothesis (EBH), die besmettelijk gapen koppelt aan emotionele overdracht, en de Attentional Bias Hypothesis (ABH), die rekening houdt met besmettelijk geeuwen als een motorische respons die onderhevig is aan verschillen in top-down aandachtsprocessen (Palagi et al., 2020).
De EBH voorspelt dat de sociale asymmetrie die wordt waargenomen in de mate van gapenbesmetting een weerspiegeling is van verschillen in de verschillende sociale binding, een proxy van emotionele binding, tussen individuen. Deze hypothese wordt ondersteund door bewijs dat de mate van besmetting met gapen een empathische trend volgt (sensu Preston en de Waal, 2002), die het hoogst is tussen individuen die een sterkste emotionele band delen. In het bijzonder vonden Norscia en Palagi (2011) dat de besmettingspercentages bij mensen het grootst zijn in reactie op verwanten en vrienden dan in reactie op kennissen en vreemden. Bij volwassen chimpansees is de geeuwbesmetting hoger tussen in-group dan bij out-group-leden (Campbell en de Waal, 2011) en bij bonobos is de geeuwbesmetting het grootst tussen individuen die meer met elkaar verbonden zijn (Demuru en Palagi, 2012). In een vergelijkend onderzoek met zowel mensen als bonobos, Palagi et al. (2014) ontdekten dat de mate van gapenbesmetting meer werd beïnvloed door de kwaliteit van de relatie tussen individuen dan door de soort waartoe de proefpersonen behoorden. Bovendien neemt de gapenbesmetting bij mensen toe met de leeftijd wanneer het vermogen om de emoties van anderen te identificeren toeneemt en afneemt met de ouderdom wanneer dat vermogen afneemt (Wiggers en van Lieshout, 1985; Anderson en Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen en Anderson, 2011; Bartholomew en Cirulli, 2014). Geeuwbesmetting neemt toe van de kindertijd tot de volwassenheid, ook bij chimpansees (Madsen en Persson, 2013).
De ABH voorspelt dat de sociale asymmetrie die wordt waargenomen bij geeuwbesmetting te wijten kan zijn aan verschillen in sociale, visuele aandacht (Massen en Gallup, 2017). In het bijzonder zouden de hoogste niveaus van besmettelijk gapen te wijten zijn aan de extra top-down, selectieve visuele aandacht die wordt besteed aan individuen die relevanter zijn voor de waarnemer, zoals bekende onderwerpen, zoals het voorkomt bij mensen en geladas, of dominanten, zoals het komt voor bij chimpansees of bonobos (Yoon en Tennie, 2010; Massen et al., 2012; Massen en Gallup, 2017).Volgens Massen en Gallup (2017) zou ABH worden ondersteund door het bestaande bewijs over de verschillende visuele detectie en visuele perceptieve codering van gezichten van bekende en / of in-groep proefpersonen in vergelijking met onbekende (bijv. Buttle en Raymond, 2003 ; Ganel en Goshen-Gottstein, 2004; Jackson en Raymond, 2006; Michel et al., 2006).
In deze studie hebben we gegevens over gapen geanalyseerd die gedurende 9 jaar zijn verzameld bij mensen in hun natuurlijke omgeving en we hebben de gevallen geëxtrapoleerd waarin de geeuw die door een proefpersoon werd uitgezonden, wel kon worden gehoord maar niet kon worden gezien door een potentiële responder (auditieve geeuw). Door alleen de gevallen te beschouwen waarin de visuele aanwijzing van de geeuwprikkel niet kon worden gedetecteerd, hebben we geverifieerd of de sociale asymmetrie die eerder werd waargenomen in de mate van geeuwbesmetting aanhield of niet. In het bijzonder hebben we de volgende alternatieve voorspellingen getest die zijn afgeleid van de twee hierboven gepresenteerde hypothesen (EBH en ABH).
Voorspelling 1a: volgens de EBH wordt de mate van geeuwbesmetting beïnvloed door de sterkte van de -individuele sociale band – een proxy van de emotionele band – per se en niet door een andere top-down, selectieve visuele aandacht die in het bijzonder aan bepaalde individuen wordt besteed. Als deze hypothese wordt ondersteund, verwachten we de sociale vooringenomenheid ook te observeren wanneer de visuele aanwijzing van de geeuwprikkel is uitgesloten en de percentages auditief besmettelijk geeuwen hoger zijn tussen sterk verbonden in vergelijking met zwak gebonden individuen.
Voorspelling 1b: volgens de ABH zouden de hogere niveaus van gapenbesmetting tussen sterk gebonden in vergelijking met zwak gebonden individuen worden gekoppeld aan de dichtstbijzijnde top-down, selectieve visuele aandacht die individuen besteden aan individuen die voor hen relevant zijn, bijv. Familie en vrienden. Als deze hypothese wordt ondersteund, zou de sociale bias die wordt waargenomen in de geeuwbesmetting moeten verdwijnen wanneer alleen auditieve geeuwen worden overwogen, omdat de visuele cue niet kan worden bijgewoond door de potentiële responder.
Materialen en methoden
Gegevensverzameling en operationele definities
Voor deze studie hebben we gekeken naar de uitgesproken geeuwen die worden uitgezonden door een onderwerp dat alleen kan worden gehoord – maar niet kan worden gezien – door een potentiële responder (hierna: auditieve geeuwen). De zender en de potentiële responders moesten zich binnen een bereik van ≤5 m bevinden. Gevocaliseerde geeuwen omvatten het gebruik van stemplooien en de geeuwen die alleen zware inspiratie / uitademing inhielden, werden niet als uitgesproken beschouwd; 294 gevallen van auditieve geeuwen werden geëxtrapoleerd uit een dataset van in totaal 2001 geeuwen verzameld over 9 jaar – van 2010 tot 2019 – door gebruik te maken van de steekproefmethode van alle voorvallen (Altmann, 1974). In het bijzonder werden auditieve geeuwen verzameld van november 2010 tot mei 2019, van 05.30 uur tot 02.30 uur, bij menselijke blanke proefpersonen, in de leeftijd van 18 tot 77 jaar, tijdens hun routinematige activiteiten, bijv. Op het werk, tijdens maaltijden, tijdens sociale bijeenkomsten, enz. ., waarbij de proefpersonen zich niet bewust waren dat ze werden geobserveerd en er geen duidelijke externe bron van angst aanwezig was. De auditieve geeuw-database omvatte 193 geeuw-potentiële responder-dyades. Afhankelijk van de situatie werd de informatie, niet genoteerd, via alfanumerieke codes geregistreerd en rechtstreeks in rekenbladen ingevoerd, in mobiele telefoons getypt of op papier geschreven, en vervolgens ingevoerd in rekenbladen voor latere uitwerking. Basisinformatie zoals leeftijd en de relatie tussen mensen was bekend bij de auteurs. De potentiële responders werden gecodeerd als in de niet-zichtbare toestand wanneer hun hoofd 180 ° werd gedraaid ten opzichte van de trigger of wanneer er een fysiek obstakel aanwezig was dat het gezichtsvermogen blokkeerde waardoor de potentiële responder het gezicht en lichaam van de trigger niet kon zien. Trigger en responder waren nooit volledig geïsoleerd (bijvoorbeeld in twee aparte kamers met gesloten deuren) van elkaar. De sociale nabijheid werd verzameld op vier niveaus: 0 = vreemden, die elkaar nog nooit hadden ontmoet; 1 = kennissen, die uitsluitend een indirecte relatie deelden op basis van een derde extern element, namelijk werkplicht (collegas) of gemeenschappelijke vrienden (vrienden van vrienden); 2 = vrienden, niet-verwante individuen die een directe relatie delen die niet uitsluitend verband houdt met een derde extern element; 3 = vaste partners en verwanten (r ≥ 0,25). Eerdere literatuur meldt dat geeuwreacties kunnen worden opgewekt binnen 5 minuten na het zien van de geeuw van iemand anders (de geeuw van de trigger) (Provine, 1986), met een maximum in de eerste minuut (Provine, 2005; Palagi et al., 2014). Literatuur meldt ook dat er vanaf de vierde minuut de grootste kans is op autocorrelatie (wat betekent dat de aanwezigheid van een geeuw uitgevoerd door een proefpersoon op t0 de kans vergroot dat dezelfde persoon nog een keer gaat geeuwen op t (0 + X) waarbij X is de toenemende tijdseenheid; Kapitány en Nielsen, 2017). Daarom hebben we rekening gehouden met de geeuwreacties die zich voordoen binnen een tijdvenster van 3 minuten vanaf de geeuw die door de trigger wordt uitgezonden.Om de autocorrelatiebias verder te verminderen, registreerden we in het geval van een ketting van geeuwen uitgezonden door de trigger (meer geeuwen uitgezonden in het tijdvenster van 3 minuten) als een reactie alleen de eerste geeuw die werd uitgevoerd na de perceptie van de laatste geeuw. We codeerden een geeuw als spontaan terwijl er geen andere proefpersoon had geeuwd in de 5 minuten voorafgaand aan de geeuw.
Statistische analyses
Voor de analyses werd rekening gehouden met de volgende variabelen: voorkomen van besmetting, gecodeerd als: 1 = aanwezigheid, 0 = afwezigheid; de sociale band werd ingevoerd met de vier hierboven gedefinieerde niveaus (0 = vreemden; 1 = kennissen; 2 = vrienden; 3 = verwanten); het geslacht van de trigger en de waarnemer werd gelabeld : M = mannelijk, F = vrouwelijk; de leeftijdsklassen van de trigger en de responder werden als volgt gecodeerd: yo = youth (18-24 jaar); ad = adult (25-64 jaar); se = senior (hierboven 65 jaar oud) (Statistics Canada, 2009); de tijdvakken waren als volgt gecodeerd: 1 = 05: 30-09: 00 uur; 2 = 09:01 uur – 12:30 uur; 3 = 12: 31-16: 00 uur; 4 = 16: 01-19: 30 uur; 5 = 19: 31-23: 00 uur; 6 = 23: 01-23: 30 uur (Giganti en Zilli, 2011). De database (zie aanvullend gegevensblad) inclusief 84 mannen, 69 vrouwen, 16 jongeren (jaar), 122 volwassenen (ad) en 15 senior (se). Om te testen of het voorkomen van geeuwen besmetting werd beïnvloed door de factoren binding (0 = vreemden; 1 = kennissen; 2 = vrienden; 3 = kin), geslacht van de trigger (Trigger_sex), geslacht van de responder (Responder_sex), de leeftijdsklasse van de trigger (Trigger_ageclass) en de leeftijdsklasse van de responder (Responder_ageclass), en tijdvak (van 1 tot 6 ), gebruikten we een gegeneraliseerd lineair gemengd model (GLMM) dat deze vijf voorspellers bevatte als vaste effecten en triggers (Trigger) en responders (Responder) identiteiten als willekeurige effecten. We hebben de modellen in R (R Core Team, 2018; versie 3.5.1) gemonteerd met de functie lmer van het R-pakket lme4 (Bates et al., 2015). We hebben de significantie van het volledige model vastgesteld door vergelijking met een nulmodel dat alleen de willekeurige effecten omvat (Forstmeier en Schielzeth, 2011). We gebruikten een likelihood ratio-test (Dobson, 2002) om deze significantie te testen (ANOVA met argument Chisq). We berekenden de p-waarden voor de individuele voorspellers op basis van likelihood ratio-tests tussen het volledige en het nulmodel met behulp van de R -functie “drop1” (Barr et al., 2013). Omdat de responsvariabele binair was, hebben we een binominale foutverdeling gebruikt. We hebben getest of de interactie tussen de seksen of de leeftijdsklassen van de trigger en de responder significant was, maar omdat ze dat niet waren, hebben we ze niet in het model opgenomen. We gebruikten een meervoudig contrastpakket (multcomp) om alle paarsgewijze vergelijkingen uit te voeren voor elk bindingsniveau met de Tukey-test (Bretz et al., 2010). We hebben de voor Bonferroni gecorrigeerde p-waarden, schatting (Est), standaardfout (SE) en z-waarden gerapporteerd.
Resultaten

Figuur 1. Lijnplot van het effect van de sociale band tussen trigger en responder (X-as) op het gemiddeld voorkomen van akoestische geeuwbesmetting (Y-as). Vrienden en verwanten vertonen significant hogere frequenties voor gapenbesmetting dan vreemden en kennissen (Tukey-test: vrienden versus vreemden p < 0,001; kin versus vreemden p < 0.001; vrienden versus kennissen p < 0.001; familie versus kennissen p < 0.001; andere combinaties, ns).
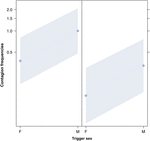
Figuur 2. Lijnplot van het effect van het geslacht van de trigger (X-as) op het gemiddelde voorkomen van akoestische geeuwbesmetting (Y-as). Effect van de triggerseks op akoestische geeuwbesmetting wanneer de responder een vrouw (rechts) en een man (links) is. De geeuw van mannen lokt meer geeuwen uit dan die van vrouwen, ongeacht het geslacht van de respondent (resultaat van de GLMM, p = 0,022). Punten vertegenwoordigen het effect dat de responsvariabele heeft op de onafhankelijke variabele geslacht van de trigger, gebaseerd op de waarde die door het model wordt voorspeld. Gekleurde banden geven het betrouwbaarheidsinterval van 95% weer.
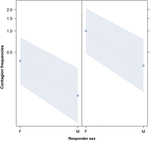
Figuur 3. Lijndiagram van het effect van het geslacht van de responder (X-as) op akoestische geeuwbesmetting gemiddeld voorkomen (Y-as). Effect van het responder geslacht op akoestische geeuwbesmetting wanneer de trigger een vrouw (rechts) of een man (links) is. Vrouwtjes reageren significant meer dan mannen, ongeacht van het geslacht van de trigger (resultaat van de GLMM, p = 0,021) Punten vertegenwoordigen het effect dat de responsvariabele heeft op de onafhankelijke variabele “geslacht van de responder”, gebaseerd op de waarde die door het model wordt voorspeld. Gekleurde banden geven het betrouwbaarheidsinterval van 95% weer.
Discussie
Deze studie toont voor de eerste keer aan dat gapenbesmetting significant wordt beïnvloed door de sociale band tussen individuen (Tabel 1), zelfs wanneer de triggerende stimuli auditieve geeuwen zijn, die we definieerden als vocaliseerde geeuwen die konden worden gehoord maar niet gezien (visuele cue niet detecteerbaar, auditieve cue detecteerbaar). Met name auditief besmettelijk geeuwen komt significant vaker voor tussen familie en vrienden dan tussen vreemden en kennissen (Figuur 1). Deze bevinding ondersteunt voorspelling 1a op basis van de EBH en niet voorspelling 1b op basis van de ABH, wat leidt tot de conclusie dat bij mensen top-down selectieve visuele aandacht niet de belangrijkste motor kan zijn van de sociale asymmetrie die wordt waargenomen bij het aantal geeuwbesmettingen (Norscia en Palagi , 2011; Norscia et al., 2016a). Ook het geslacht van de trigger en het geslacht van de ontvanger hadden een significant effect op de mate van gapenbesmetting, waarbij mannen – als triggers – vaker door anderen werden gereageerd dan vrouwen (figuur 2) en vrouwen vaker op andermans geeuwen reageerden dan mannen (Figuur 3).
In tegenstelling tot Bartholomew en Cirulli (2014), vonden we geen leeftijdseffect op geeuwbesmetting, hoogstwaarschijnlijk omdat onze database over auditieve geeuwen een sterke prevalentie had onder volwassenen (25-64 jaar oud). De hoogste niveaus van auditieve geeuwbesmetting bij vrouwen in vergelijking met mannen bevestigen de gendervooroordelen die worden waargenomen in naturalistische omstandigheden bij mensen die vatbaar zijn voor geeuwbesmetting door Norscia et al. (2016a, b) bij het overwegen van een grotere dataset die ook geeuwen omvatte die door de potentiële responder konden worden gezien (met een gapende sensorische modaliteit – zien, horen of beide – die de respons niet beïnvloedden). De gendervooroordelen zijn ook gedeeltelijk in overeenstemming met eerdere resultaten die zijn verkregen in gecontroleerde omgevingen, inclusief de visuele cue (Chan en Tseng, 2017; maar zie Norscia en Palagi, 2011; Bartholomew en Cirulli, 2014). De hypothese is dat de hoge mate van gapenbesmetting bij vrouwen emotionele besmetting zou kunnen veroorzaken (Norscia et al., 2016a), in het licht van naar verluidt hogere empathische vermogens – gerelateerd aan moederschap – van vrouwen in vergelijking met mannen (Christov-Moore et al. ., 2014). Deze kwestie staat echter nog steeds ter discussie omdat culturele verschillen tussen menselijke samenlevingen de dynamiek van sociale binding op een andere manier kunnen vormen. In dit kennisstadium is het daarom ingewikkeld om culturele factoren, de kwaliteit van interpersoonlijke relaties en genderinvloed bij de verspreiding van geeuwbesmetting te ontwarren. Onze resultaten laten ook zien dat mannen beter presteren dan vrouwen als triggers en de meest spaarzame hypothese hiervoor zou kunnen zijn dat de vocalisaties van mannen beter gehoord kunnen worden in natuurlijke omgevingen, vaak gekenmerkt door achtergrondgeluiden. De perceptie van het geslacht van de stem berust in de eerste plaats op de fundamentele frequentie die gemiddeld een octaaf lager is bij mannenstemmen dan bij vrouwenstemmen, waarbij vocalisaties met een lagere frequentie verder reiken dan die met hoge frequenties (Marten en Marler, 1977; Latinus en Taylor, 2012) . Voor zover wij weten, is er echter geen specifiek onderzoek naar de mogelijke gendervooroordelen in hoorbaarheid van geeuwen en verder onderzoek met experimentele proeven in gecontroleerde toestand is daarom nodig om deze speculatie te verifiëren.
In dit onderzoek hebben we ook gevonden dat de verschillen in geeuwbesmetting tussen categorieën (familie en vrienden, vreemden en kennissen) niet kunnen worden verklaard door verschillen in top-down, selectieve visuele aandacht. Deze bevinding is in lijn met eerdere literatuur. Bij besmettelijk gapen lijken hersengebieden te zijn betrokken die meer verband houden met het oriënterende bottom-up netwerk dan met top-down gerelateerde gebieden. Bovendien is gapenbesmetting niet gevoelig voor de sensorische signalen die in het signaal aanwezig zijn (auditief, visueel of audiovisueel) (Arnott et al., 2009; Norscia en Palagi, 2011), noch beïnvloed door het visuele perspectief van de triggerende stimulus ( geeuwen in oriëntaties van 90 °, 180 ° en 270 ° kunnen gaapreacties opwekken als frontale, 0 ° geeuwen; Provine, 1989, 1996). Chan en Tseng (2017) ontdekten dat het vermogen om een geeuw als zodanig te detecteren (perceptuele detectiegevoeligheid) gerelateerd was aan de duur van het kijken naar de ogen van het gezicht dat de stimulus vrijgeeft, maar oog-blikpatronen waren niet in staat om besmettelijk geeuwen te moduleren. Bij chimpansees waren de besmettelijke geeuwfrequenties het hoogst tussen individuen uit dezelfde groep dan tussen individuen uit verschillende groepen, zelfs als de respondenten langer naar chimpanseevideos uit de outgroep keken (Campbell en de Waal, 2011). Het argument dat visuele selectieve aandacht de besmettingspercentages van de geeuw in een bepaalde richting kan beïnvloeden (proefpersonen reageren meer op familie dan vreemden) wordt ook ondermijnd door de afwezigheid van een specifiek patroon van sociale aandacht bij menselijke en niet-menselijke primaten. Via eye-tracking (toegepast om de kijktijd te meten) en door onbekende gezichten te laten zien aan hun proefpersonen, Méary et al.(2014) merkten op dat mensen scheef stonden in de richting van gezichten van hun eigen ras, terwijl de aandacht van resusapen meer werd aangetrokken door nieuwe dan door gezichten van dezelfde soort. Kawakami et al. (2014) merkten op dat proefpersonen meer aandacht besteedden aan de ogen van etnische groepsleden en aan neus en mond van etnische outgroepsleden. Dezelfde studie onthulde ook dat visuele aandacht niet afhankelijk was van het doelwitras. Door te meten hoe lang de proefpersonen naar het scherm staarden, staarden Whitehouse et al. (2016) merkten op dat Berberapen meer aandacht besteedden aan het krabben van videos van niet-vreemden dan aan vreemde individuen, maar merkten ook op dat binnen de niet-vreemden, makaken de meeste aandacht besteedden aan die individuen waarmee ze een zwakke sociale band deelden. Schino en Sciarretta (2016) hebben door middel van het meten van de snelheden waargenomen dat mandrillen meer naar hun eigen verwanten keken dan naar niet-verwanten, maar ook meer naar dominant dan naar ondergeschikte groepsgenoten. Daarom beschrijven deze onderzoeken (gebruikt om ABH te ondersteunen) geen enkel patroon van selectieve aandacht. Een ander belangrijk punt om te overwegen is de definitie van vertrouwdheid en groepslidmaatschap die in de meeste onderzoeken wordt gebruikt om ABH te ondersteunen (Massen en Gallup, 2017). Deze onderzoeken lieten een betere visuele detectie en visuele perceptieve codering zien van gezichten van bekende / in-groep proefpersonen in vergelijking met onbekende, maar definieerden vertrouwdheid en groepslidmaatschap niet op basis van de persoonlijke relaties tussen individuen. In plaats daarvan werd bekendheid of groepslidmaatschap gedefinieerd op grond van indirecte kennis (bijv. Foto van beroemde mensen of van een onderwerp dat al in vooronderzoeken werd getoond) of algemeen ras (bijv. Buttle en Raymond, 2003; Ganel en Goshen- Gottstein, 2004; Jackson en Raymond, 2006; Michel et al., 2006). Deze definitie is prima voor de doeleinden van deze onderzoeken, maar het is niet zo erg als de resultaten worden gebruikt om alternatieve verklaringen voor te stellen voor de invloed die echte sociale binding – gebaseerd op echte relaties – kan hebben op een fenomeen, in dit geval geeuwbesmetting. . Michel et al. (2006) merkten op dat blanke en Aziatische proefpersonen gezichten van hetzelfde ras beter konden herkennen, maar dit verschil was niet aanwezig bij Aziatische proefpersonen die al ongeveer een jaar onder blanken woonden. Een ander punt van discussie betreft de aanwezigheid van gapenbesmetting bij kinderen met een autismespectrumstoornis (ASS), die vaak veranderingen in visuele aandacht laten zien (Richard en Lajiness-O’Neill, 2015). Bij ASS-kinderen kan gapenbesmetting afwezig zijn (Senju et al., 2007), verminderd (Helt et al., 2010), of vergelijkbaar met typisch ontwikkelende kinderen wanneer de proefpersonen worden aangezet om hun aandacht te verleggen naar de videostimulus tijdens de experimentele proeven. (Usui et al., 2013). In een recente studie hebben Mariscal et al. (2019) ontdekten dat gapenbesmetting bij ASS-kinderen positief verband hield met de bloedconcentratie van oxytocine, het hormoon dat betrokken is bij ouderlijke en sociale gehechtheid (Decety et al., 2016) en stelde dat gapenbesmetting bij ASS-kinderen mogelijk verband houdt met een variabel gemiddelde. oxytocineconcentraties in verschillende studiecohorten (Mariscal et al., 2019). Deze bevinding is in lijn met de EBH-hypothese die de kans op geeuwbesmetting koppelt aan sociale binding, wat een weerspiegeling kan zijn van emotionele binding.
Conclusie
Onze studie draagt bij aan de discussie over de mechanismen die ten grondslag liggen aan de sociale asymmetrie bij geeuwbesmetting (voor een kritische recensie: zie Adriaense et al., 2020; Palagi et al., 2020), door aan te tonen dat geeuwbesmetting waarschijnlijk wordt geassocieerd met bottom-up, in plaats van met top-down, selectieve aandacht. Bottom-up-aandacht wordt voornamelijk geleid door de sensorische perceptie van de uitlokkende stimulus, terwijl top-down, selectieve aandacht een vrijwillig, aanhoudend proces is waarin een bepaald item intern wordt geselecteerd en erop wordt gefocust of onderzocht (Katsuki en Constantinidis, 2014). In dit opzicht was de akoestische stimulus (auditieve geeuw) die door de trigger werd uitgezonden, gehoord en kon een geeuwreactie in de ontvanger opwekken, ook al schonk de ontvanger geen vrijwillige visuele aandacht aan de trigger. Bovendien waren de responspercentages bij geeuwen sociaal gemoduleerd, waarbij de auditieve geeuwbesmetting het hoogst was bij individuen die het sterkst met elkaar verbonden waren. Daarom is selectieve aandacht van bovenaf niet de belangrijkste motor van de sociale asymmetrie die wordt waargenomen bij geeuwbesmetting, wat een door stimuli aangedreven fenomeen lijkt te zijn dat verband houdt met aandachtsprocessen van onderaf. Verder onderzoek is nodig om te begrijpen of en op welke manier andere vormen van aandacht of pre-attente stadia geeuwbesmetting kunnen beïnvloeden.
Verklaring gegevensbeschikbaarheid
De dataset die voor dit onderzoek is gebruikt wordt verstrekt in het aanvullende materiaal.
Ethische verklaring
De studie met menselijke deelnemers werd beoordeeld en goedgekeurd door Comitato di Bioetica dAteneo (University Bioethical Committee) —University of Turin (ref nr. 451945).Schriftelijke geïnformeerde toestemming voor deelname was niet vereist voor deze studie in overeenstemming met de nationale wetgeving en de institutionele vereisten.
Bijdragen van auteurs
IN en EP hebben gegevens verzameld en de manuscript. AZ hielp met het verzamelen van gegevens en revisie van manuscripten, cijfers en tabellen. MG voerde statistische analyses uit en schreef het gerelateerde deel van het manuscript.
Belangenconflict
De auteurs verklaren dat het onderzoek is uitgevoerd zonder dat er commerciële of financiële relaties waren die worden opgevat als een mogelijk belangenconflict.
Dankbetuigingen
De auteurs willen prof. Roberto Barbuti en prof. Cristina Giacoma bedanken voor hun steun aan het onderzoek aan de Universiteit van Pisa en de Universiteit van Turijn.
Aanvullend materiaal
Altmann, J. (1974). Observationeel onderzoek naar gedrag: steekproefmethoden. Gedrag 49, 227-266. doi: 10.1163 / 156853974×00534
PubMed Abstract | CrossRef Volledige tekst | Google Scholar
Anderson, J. R., en Meno, P. (2003). Psychologische invloeden op gapen bij kinderen. Curr. Psychol. Lett. 11, 1-7.
Google Scholar
Bartholomew, A. J., en Cirulli, E. T. (2014). Individuele variatie in vatbaarheid voor besmettelijke geeuwen is zeer stabiel en grotendeels onverklaard door empathie of andere bekende factoren. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | CrossRef Volledige tekst | Google Scholar
Bretz, F., Hothorn, T., en Westfall, P. (2010). Meerdere vergelijkingen met R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Ethologische studie van gapen bij primaten. I. Kwantitatieve analyse en studie van het oorzakelijk verband bij twee soorten apen uit de Oude Wereld (Cercocebus albigena en Macaca fascicularis). Ethology 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef volledige tekst | Google Scholar
Dobson, A. J. (2002). Een inleiding tot gegeneraliseerde lineaire modellen, 2e ed. Boca Raton, FL: Chapman en Hall.
Google Scholar
Katsuki, F., en Constantinidis, C . (2014). Bottom-up en top-down aandacht: verschillende processen en overlappende neurale systemen. Neurowetenschapper 20, 509-521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef Volledige tekst | Google Scholar
Singer, T. (2006). De neuronale basis en ontogenie van empathie en gedachtenlezen: literatuuroverzicht en implicaties voor toekomstig onderzoek. Neurosci. Biobehav. Rev. 30, 855-863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | CrossRef Volledige tekst | Google Scholar
Statistics Canada (2009). Leeftijdscategorieën, levenscyclusgroeperingen. Ottawa: Statistics Canada.
Google Scholar