Informatieverwerking is essentieel in alle wetenschapsgebieden. In de moleculaire biologie is het centrale dogma, voor het eerst bedacht door Francis Crick (Crick, 1958, 1970), een klassieke ruggengraat van levende cellen om fundamenteel processen uit te voeren van celdeling tot de dood via de DNA-, RNA- en eiwitinformatiepaden. Meer specifiek beschrijft het centrale dogma de overdracht van sequentie-informatie tijdens DNA-replicatie, transcriptie naar RNA en vertaling in aminozuurketens die eiwitten vormen. Tegelijkertijd stelt het ook dat informatie niet van eiwit naar eiwit of nucleïnezuur kan stromen.
Sinds de opkomst van systemische en high-throughput-benaderingen in de afgelopen twee decennia, zijn deze brede stappen, die niet omvatten complexe regelgevingsdetails zijn onder de loep genomen. De ontbrekende regulerende kenmerken, zoals de mechanismen voor het corrigeren / herstellen van DNA en alternatieve splitsing van pre-mRNA, introduceren verschillende tussenstappen. Deze extra stappen verstoren de belangrijkste stappen van het dogma en veranderen waarschijnlijk de informatiedynamiek. Bovendien lijken epigenetica, of de rol die wordt gespeeld door chromatinestructuren, DNA-methylatie en histonmodificaties, ook in te gaan tegen de eenvoudige paden van het dogma (Shapiro, 2009; Luco et al., 2011). Eiwitsplitsing, of het vermogen van een eiwit (inteïnen) om zijn eigen sequentie te veranderen, recent ontdekt (Volkmann en Mootz, 2012) en prionen, die andere eiwitsequenties modificeren (Prusiner, 1998), omzeilen de informatieoverdrachtsroute van de dogma. Andere onderzoeken meldden fouten of mismatches tussen RNA-sequenties en hun coderende DNA (Hayden, 2011; Li et al., 2011). Samengevat werpen deze gegevens twijfels op over de geldigheid van het centrale dogma in de context van de huidige wetenschap, en daarom vragen ze de eenvoud van lineaire informatiestroom (DNA naar RNA en RNA naar eiwit).
Om de zaken in perspectief te plaatsen, hebben we analytische hulpmiddelen nodig die de zorgen of discrepanties met betrekking tot de al lang bestaande theorie onderzoeken. Een eenvoudige, maar zeer nuttige techniek voor het zoeken naar globale eigenschappen in datasets met hoge doorvoer is statistische correlatieanalyse, die op grote schaal en met succes is gebruikt om patronen in complexe systemen zoals het weer te observeren (Stewart, 1990), aandelenmarkten (Lo en MacKinlay). , 1988) en kosmologie (Amati et al., 2008). Er zijn verschillende soorten correlatieanalyses die zowel lineaire (bijv. Pearson-product-moment) als niet-lineaire (bijv. Spearmans rang, wederzijdse informatie) afhankelijkheden evalueren (Steuer et al., 2002; Rosner, 2011). is de Pearson product-moment correlatieanalyse het populairst geworden vanwege het vermogen om de organisatiestructuur in de eenvoudigste vorm weer te geven.
In de biologie zijn er talloze werken geweest die de correlaties in het mRNA en eiwitexpressiegegevens (zie hieronder en tabel 1). In theorie, wanneer twee monsters met hoogdimensionale (zoals microarray en proteomische) gegevens worden vergeleken, geven de correlatieanalyses een maat voor afwijking van eenheid als bron van verschil tussen de monsters . In het kort, twee steekproeven met identieke en volledig niet-identieke informatie zullen respectievelijk de eenheid (R2 = 1) en nul (R2 = 0) correlatie tonen.
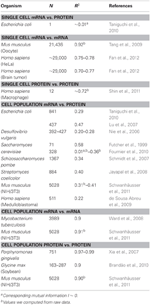
Tabel 1. mRNA en proteïne-expr essiecorrelaties in verschillende organismen.
Perfecte correlatie (R2 = 1) is een geïdealiseerde situatie die verre van werkelijkheid is, aangezien technische of experimentele ruis alleen interfereert en vermindert de correlatie. Bovendien hebben de afgelopen jaren het bestaan van biologische ruis aan het licht gebracht: de studies van individuele cellen en moleculen hebben stochasticiteit aangetoond in de dynamiek van genexpressie als gevolg van het combinatorische effect van lage moleculaire kopieaantallen en de kwantiteit van de dynamica van de promotor (Raj en van Oudenaarden, 2009; Eldar en Elowitz, 2010). Aan de andere kant vertonen klonale celpopulaties heterogeniteit in de niveaus van een bepaalde eiwitexpressie per cel op elk gemeten tijdstip (Chang et al., 2008). Samen zijn stochasticiteit en heterogeniteit essentieel voor het produceren van diversificatie van het lot van cellen, fenotypische variaties en versterking van intracellulaire signalen (Locke et al., 2011; Selvarajoo, 2012).
De stochastische fluctuaties, of intrinsieke ruis, veroorzaken de expressie van een moleculaire soort varieert in tijd en tussen cellen, wat leidt tot niet-gecorreleerde reacties (Elowitz et al., 2002). Dit is vooral prominent voor mRNAs en eiwitten met een laag aantal kopieën. Zo kan de correlatie tussen monsters (cellen) worden verlaagd vanwege intrinsieke ruis (Figuur 1A). Andere bronnen van biologische ruis als gevolg van extrinsieke factoren zijn onder meer variabiliteit in celgrootte, aantal moleculaire kopieën en omgevingsfluctuaties tussen individuele cellen.Deze factoren verstoren het deterministische centrale dogma en veranderen waarschijnlijk sterke correlaties in zwakkere (Figuur 1B).
Een recente studie vergeleek Escherichia coli-mRNA en eiwitexpressies tussen individuele cellen op het niveau van één molecuul en leverde een scenario op dat het centrale dogma diep in twijfel trekt. Taniguchi et al. (2010) onthulden dat er geen correlatie (R2 ~ 0) is tussen individuele tufA-mRNA- en eiwitniveaus in afzonderlijke cellen. Ze concludeerden met name dat het gebrek aan correlatie waarschijnlijk te wijten is aan verschillen in mRNA- en eiwitlevensduur. Hoewel dit een plausibele verklaring is, stellen Taniguchi et al. waren voorzichtig om de langdurige hypothese niet te weerleggen door te beweren dat tijdgemiddelden van mRNA-niveaus zouden moeten correleren met eiwitniveaus. Er was echter geen bewijs om aan te tonen dat dit het werkelijke geval is, en toen we niet-lineaire afhankelijkheden evalueerden met behulp van wederzijdse informatie (Steuer et al., 2002; Tsuchiya et al., 2010) in Taniguchi et al. dataset, vonden we dat het resultaat niet-afhankelijk was, d.w.z. I ~ 0. Dit bevestigt dat mRNA naar eiwitexpressies tussen individuele cellen op het niveau van één molecuul duidelijk geen verband houden. Bovendien is het bij inzoomen op het niveau van een enkel molecuul in de correlatiegrafiek duidelijk dat hun paarsgewijze correlaties zwak zijn (Figuur 1A, insert, ter illustratie).
Met name op celpopulatieniveau, Taniguchi et al. waren in staat om een relatief hoge correlatie tussen mRNA en eiwitexpressies te laten zien met R2 = 0,29 (Figuur 2A). In feite is een andere onafhankelijke studie van Lu et al. (2007), voor E. coli-populatie, vertoonden ook een relatief hoge correlatie (R2 = 0,47). Vergelijkbare analyses uitgevoerd op Saccharomyces cerevisiae (Futcher et al., 1999), NIH / 3T3-fibroblasten van muizen (Schwanhäusser et al., 2011) en verschillende andere celpopulaties (Nie et al., 2006; Schmidt et al., 2007; Jayapal et al., 2007; al., 2008; de Sousa Abreu et al., 2009) vertoonden allemaal gecorreleerde structuren tussen transcriptoom-brede en proteoom-brede expressies (tabel 1). Dus waarom is er geen verband tussen individuele mRNA- en eiwitexpressies in afzonderlijke cellen, terwijl er op populatieniveau collectieve relaties worden waargenomen tussen grootschalige mRNA- en eiwitexpressies?
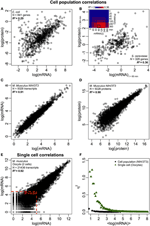
Figuur 2. Omics-brede expressiecorrelaties. Celpopulaties: mRNA-eiwitcorrelaties in (A) E. coli (Taniguchi et al., 2010) en (B) S. cerevisiae (Fournier et al., 2010) tussen mRNA-expressies op t = 60 min en eiwitexpressies op t = 360 min. Invoegen: correlatiematrix tussen alle tijdstippen toont een vertraagde toename van correlaties tussen mRNA en eiwitten. (C) mRNA- en (D) eiwitexpressies tussen twee monsters van muizen NIH / 3T3-cellen (Schwanhäusser et al., 2011). Enkele cellen: (E) mRNA-expressies tussen twee eicellen (Tang et al., 2009). De rode stippellijnen geven de gebieden aan met lage mRNA-expressies (log (mRNA) < 5). (F) Ruis (η2) versus log (mRNA-expressies) voor celpopulatie (NIH / 3T3, zwarte stippen, Schwanhäusser et al., 2011) en enkele cellen (eicellen, groene driehoeken, Tang et al., 2009). Elke stip vertegenwoordigt de waarde voor een groep van P = 100 mRNAs. η2 is bijna nul voor de celpopulatie voor alle mRNA-expressies. Voor enkele cellen is η2 het hoogst voor mRNAs met de laagste kopieaantallen, en nadert nul voor hogere kopieaantallen.
Wij denken er zijn twee belangrijke redenen voor de verschillen. Ten eerste, zoals eerder opgemerkt, vermindert ruis, of dit nu biologisch of niet-biologisch van aard is, de correlatie. Omdat analyses op enkele cellen het belang van stochasticiteit en variabiliteit hebben aangetoond, zijn deze effecten cruciaal voor het verminderen van de correlaties van eencellige. Op ensemble-niveau, wanneer cellen worden bemonsterd in een populatie, wordt de totale (intrinsieke + extrinsieke) ruis verminderd, aangezien willekeurige ruis wordt onderdrukt over alle moleculaire uitdrukkingen (figuren 1C-F), om een gemiddelde respons en zelforganisatie te onthullen ( Karsenti, 2008; Selvarajoo, 2011; Hekstra en Leibler, 2012; Selvarajoo en Giuliani, 2012). Daarom komt er een goede mate van correlatie tussen mRNA-proteïne-expressie naar voren. Ten tweede werd voor de eencellige studie (Taniguchi et al., 2010) de individuele mRNA-eiwitexpressiecorrelatie vergeleken over talrijke cellen. In celpopulatiestudies wordt de vergelijking echter in zijn geheel gemaakt, over duizenden mRNAs en eiwitten over verschillende orden van grootte groter dan het bereik van expressie dat wordt gevonden voor één molecuul tussen cellen. Dit leidt daarom tot hogere correlaties op populatieniveau aangezien het effect van enkele moleculaire variaties verwaarloosbaar wordt.
Ondanks dat gecorreleerde structuren worden waargenomen voor celpopulaties, zijn er tastbare redenen voor de grote afwijking van perfecte correlatie.Zoals eerder opgemerkt, is een belangrijk punt dat mRNAs en eiwitten opeenvolgend worden gelokaliseerd met verschillende ontbrekende processen, niet vertegenwoordigd in het centrale dogma. Het toevoegen van de ontbrekende tussenproducten langs een biochemische route zal een merkbare vertraging in de informatiestroom veroorzaken (Selvarajoo, 2006, 2011; Piras et al., 2011), en de correlatie daartussen zou als gevolg daarvan kunnen lijden. Dit zou ook deel kunnen uitmaken van het feit dat is opgemerkt door Taniguchi et al. dat mRNA- en eiwitexpressies verschillende levensduren hebben. Deze stelling wordt met name ondersteund in een recent werk over S. cerevisiae behandeld met Rapamycin dat aantoonde dat de temporele correlaties van mRNA-eiwitexpressie aanvankelijk laag waren, R2 = 0,01 na 40 minuten, niettemin, meer dan 360 minuten na verstoring, nam de correlatie toe, R2 = 0,36 (Fournier et al., 2010, Figuur 2B). De gegevens geven aan dat bij chemische verstoring de initiële respons tussen mRNA- en eiwitexpressies afwijkt als gevolg van tijdsvertraging en verschillende kinetische mechanismen daartussen, evenals secundaire effecten zoals autocriene of paracriene signaalinterferentie (Shvartsman et al., 2002; Isalan et al., 2008). Wanneer de effecten van de verstoring in de loop van de tijd worden afgezwakt, treedt herstel van correlaties op.
Om de veronderstelling dat opeenvolgende vertragingsprocessen of verschillende levensduren cruciaal zijn voor het verminderen van mRNA-eiwitcorrelaties verder te controleren, vergeleken we R2 tussen de dezelfde moleculaire soort van het centrale dogma (bijv. tussen mRNA en mRNA) in celpopulaties en enkele cellen. De transcriptoom-brede mRNA-mRNA-expressiecorrelatie tussen replicaten van NIH / 3T3 (Schwanhäusser et al., 2011) (Figuur 2C) en Mycobacterium tuberculosis (Ward et al., 2008) celpopulatiemonsters zijn beide erg hoog, met R2 > 0.9 (tabel 1). Dergelijke sterke correlaties worden ook waargenomen tussen populatiemonsters voor eiwit-eiwit-expressies in NIH / 3T3-cellen (Schwanhäusser et al., 2011) (Figuur 2D), Porphyromonas gingivalis (Xia et al., 2007) en Glycine max (Brandão et al. , 2010) (tabel 1). Aangezien deze gegevens, die dezelfde soorten vergelijken, zeer hoge correlaties opleveren, is het denkbaar dat de opeenvolgende vertragingsprocessen of verschillende levensduren verantwoordelijk zijn voor het verlagen van de correlatiestructuren op populatieniveau tussen mRNA en eiwitexpressies.
In enkele muiseicellen ( Tang et al., 2009), wanneer volledige mRNA-mRNA-expressies worden vergeleken, wordt een sterk gecorreleerde structuur waargenomen (R2 = 0.92, Figuur 2E). Als u zich echter alleen richt op laag tot expressie gebrachte mRNAs (met logaritmische uitdrukkingen < 5), verlaagt de stochastische ruis de paarsgewijze correlatie behoorlijk dramatisch (R2 < 0,54). Om dit resultaat te onderzoeken, hebben we ruis geëvalueerd, η2 = σ2XY / μ2XY, over hele mRNA-expressies (Figuur 2F). We merkten op dat η2 het hoogst is voor de laagste expressies, vanwege het uitgesproken effect van stochastische fluctuaties in vergelijking met hun expressies, en benadert nul voor hogere expressies, waar dergelijke ruis minder significant wordt (Piras et al., 2012). Voor celpopulatie wordt, zoals verwacht, bijna geen ruis waargenomen over het gehele expressiebereik als gevolg van het onderdrukken van willekeurige ruis (figuren 1E, F).
Sterk gecorreleerde structuren voor volledige mRNA-mRNA-expressies waren ook gerapporteerd voor een enkele kankercel (Fan et al., 2012), zij het minder significant met R2 ~ 0,7 (Tabel 1). Bovendien vertoonden de vergelijking van proteïne-proteïne-expressies in door LPS gestimuleerde menselijke macrofagen ook hoge correlaties, R2 ~ 0,72 (Shin et al., 2011) (Tabel 1). Hoewel er geen correlatie is tussen individuele mRNA-eiwitexpressies in enkele cellen, is de grootschalige of omics-brede correlatie tussen dezelfde moleculaire soorten in enkele cellen erg hoog.
Dus of het nu gaat om enkele cellen of celpopulaties. geven de omics-brede gegevens aan dat de correlaties tussen dezelfde moleculaire soort (mRNA versus mRNA, en eiwit versus eiwit) merkbaar hoger zijn dan tussen verschillende soorten (mRNA versus eiwit). Dit weerspiegelt het feit dat, hoewel tijdvertragingsprocessen en verschillende levensduren de sleutel zijn voor het verminderen van correlaties, deze mechanismen niet voldoende zijn om het gebrek aan correlatiestructuur te ondersteunen die wordt waargenomen tussen individuele transcripten van afzonderlijke cellen naar eiwitexpressies.
Dus door grootschalige expressie van mRNAs en eiwitten van verschillende cellulaire systemen te onderzoeken, hebben we aangetoond dat correlatiestructuren op mondiale schaal ontstaan. De correlatieanalyses onthullen echter alleen de connectiviteit tussen twee geteste monsters, en niet de richting van informatiestroom. Wil het centrale dogma geldig zijn op mondiale schaal, dan moet de algehele informatiestroom van DNA naar eiwitten zijn. Een dergelijke informatiestroom is aangetoond door talloze andere onderzoeken waarbij de receptoren van celpopulaties worden verstoord en de resulterende dynamiek van transcriptiefactoren die binden aan DNA en de inductie van grootschalige genexpressies (Figuur 3A).In het geval van LPS-gestimuleerde immuuncellen is bijvoorbeeld aangetoond dat de activering van de transcriptiefactor NF-KB plaatsvindt na ongeveer 15 minuten (Liu et al., 1999), de inductie van de stroomafwaartse genen op ongeveer 30 minuten. min (Liu et al., 1999; Xaus et al., 2000; Selvarajoo et al., 2008), en de vertaling van de overeenkomstige eiwitten in het gebied van 60-90 min (Kawai et al., 1999; Xaus et al. ., 2000) (Figuur 3B). Een dergelijke opeenvolgende richting van de totale informatiestroom van transcriptie naar vertaling wordt ook waargenomen voor bacteriële systemen, zoals E. coli, op celpopulatieniveau (Golding et al., 2005).

Figuur 3. De informatiestroom van centraal dogma. (A) Schematische voorstelling van LPS / TLR4-geïnduceerde TNF-expressie, via transcriptiefactor NF-KB en tnf-gen, na lineaire informatiestroom. (B) Experimentele temporele profielen van promotorbindingsactiviteit van NF-KB (bovenste panelen), tnf (middelste panelen) en TNF (onderste panelen) expressies op celpopulatieniveau. (C) Schematische temporele profielen van promotordynamica, mRNA en eiwitexpressies op eencellig niveau (Raj en van Oudenaarden, 2009).
Als alternatief onthullen onderzoeken met eencellige resolutie willekeurige fluctuaties in de lineaire informatiestroom: de transcriptiefactoren die binden aan DNA-promotorregios zijn kwantitatief, resulterend in barstgedrag van de mRNA-transcriptie en vervolgens variabiliteit in de eiwittranslatie, zelfs tussen identieke cellen (Figuur 3C) (Raj en van Oudenaarden, 2009; Eldar en Elowitz, 2010; Locke et al., 2011; Hekstra en Leibler, 2012; Selvarajoo, 2012). Als gevolg hiervan is de individuele moleculaire respons voor afzonderlijke cellen op een bepaald tijdstip nogal luidruchtig vergeleken met de gemiddelde populatieschaal (Selvarajoo, 2011).
Conclusies
De voorbeelden in dit artikel belicht de verschillen in de volgorde van correlatiewaarden die zijn waargenomen tussen soorten in het centrale dogma over celpopulaties en enkele cellen. De statistische analyses van celpopulaties schetsen een beeld dat de expressiecorrelatie tussen dezelfde moleculaire soorten erg hoog is en tussen soorten matig hoog. Hoewel eencellige correlaties tussen dezelfde soort vergelijkbaar zijn met celpopulaties, vertoonden ze een grotere spreiding in hun expressiegrafieken vanwege het uitgesproken effect van biologische ruis, vooral voor transcripten met lage kopieaantallen. Met name de paarsgewijze correlatie van de afzonderlijke cellen wordt nul voor individuele moleculen (Taniguchi et al., 2010). In feite is bekend dat stochastische fluctuaties en variabiliteit in moleculaire expressies functioneel zijn bij het genereren van beslissingen over het lot van de cel en het kantelen van cellulaire toestanden (Losick en Desplan, 2008; Eldar en Elowitz, 2010; Kuwahara en Schwartz, 2012). Wij geloven dat de sterke omics-brede correlaties optreden als gevolg van nauwe gen- en eiwitregulerende netwerken over duizenden moleculen (Barabási en Oltvai, 2004; Karsenti, , 2008) resulterend in opkomende gemiddelde reacties. Bij het analyseren van een klein aantal of individuele moleculen kan de correlatiestructuur niet worden waargenomen.
Over het algemeen is het denkbaar dat het bekijken van de informatiestroom van enkel DNA naar eiwit vraagtekens zet bij het centrale dogma aangezien de respons van elk molecuul op een bepaald moment waarschijnlijk niet zal correleren. Wereldwijd suggereert de waarneming van de gemiddelde deterministische respons echter dat het netto evenwicht van het genet ic-informatie blijft uiterst rechts van de paden. Daarom moet het centrale dogma worden gezien als een macroscopische cellulaire informatiestroom op omics-brede schaal, en niet als een enkel gen naar eiwitniveau. Als zodanig geloven we dat de eenvoud ervan een van de meest invloedrijke theoretische pijlers van levende systemen zal blijven.
Verklaring over belangenconflicten
De auteurs verklaren dat het onderzoek is uitgevoerd in de afwezigheid van enige commerciële of financiële relaties die kunnen worden opgevat als een potentieel belangenconflict.
Dankbetuigingen
Kentaro Hayashi wordt bedankt voor zijn opmerkingen. Het onderzoeksfonds van de stad Tsuruoka en de prefectuur Yamagata wordt gewaardeerd voor hun steun.
Crick, F. (1958). Over eiwitsynthese. Symp. Soc. Exp. Biol. 12, 139–163.
Pubmed Abstract | Pubmed Full Text
Crick, F. (1970). Centraal dogma van moleculaire biologie. Nature 227, 561-563.
Pubmed Abstract | Pubmed Full Text
Hayden, E. C. (2011). Bewijs van veranderd RNA roept discussie op. Nature 473, 432.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Full Text
Hekstra, D. R., en Leibler, S. (2012). Contingentie en statistische wetten in replicerende microbiële gesloten ecosystemen. Cell 149, 1164-1173.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Kuwahara, H., en Schwartz, R. (2012).Stochastische steady-state-winst in een genexpressieproces met mRNA-degradatiecontrole. J. R. Soc. Interface 9, 1589-1598.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Nie, L., Wu, G., en Zhang, W. (2006). Correlatie van mRNA-expressie en eiwitovervloed beïnvloed door meerdere sequentiekenmerken gerelateerd aan translationele efficiëntie in Desulfovibrio vulgaris: een kwantitatieve analyse. Genetics 174, 2229-2243.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Prusiner, S. B. (1998). Prionen. Proc. Natl. Acad. Sci. U.S.A. 95, 13363–13383.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Rosner, B. (2011). Fundamentals of Biostatistics. 7e Edn. Boston, MA: Duxbury Press.
Selvarajoo, K. (2006). Het ontdekken van differentiële activeringsmechanismen van de tolachtige receptor 4-signaalroutes in MyD88-knockouts. FEBS Lett. 580, 1457–1464.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Selvarajoo, K. (2011). Macroscopische behoudswet onthuld in de populatiedynamiek van tolachtige receptorsignalering. Cell Commun. Signaal. 9, 9.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Selvarajoo, K. (2012). Inzicht in multimodale biologische beslissingen op basis van eencellige en populatiedynamiek. Wiley Interdiscip. Rev. Syst. Biol. Med. 4, 385-399.
Pubmed Abstract | Pubmed volledige tekst | CrossRef Volledige tekst
Stewart, T. R. (1990). Een decompositie van de correlatiecoëfficiënt en het gebruik ervan bij het analyseren van voorspellingsvaardigheden. Weervoorspelling. 5, 661-666.