Introduksjon
Gjesping er en ufrivillig sekvens av munnåpning, dyp inspirasjon, kort apné og mer eller mindre langsom utløp (Baenninger, 1997; Walusinski og Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Når man fremkaller et gjesp, kan det ikke undertrykkes totalt. Derfor har det blitt definert som et stereotype eller reflekslignende mønster (Lehmann, 1979; Provine, 1986). I Homo sapiens har flere hypoteser blitt fremført med variabel støtte for å forklare mekanismer og funksjoner av spontan gjesp, som oksygenering (respiratorisk funksjon forårsaket av hypoksi), stressrelatert oppførsel (forårsaket av opphisselse) eller termoregulering (forårsaket av hypertermi Guggisberg et al., 2010; Massen et al., 2014; Gallup and Gallup, 2019). Å være en fysiologisk respons, kan gjesping påvirkes av interne og eksterne faktorer som tiden på dagen (Giganti og Zilli, 2011) eller intrakraniell / hjernetemperatur (Gallup og Eldakar, 2013).
Gabe kan være selvstyrt og / eller vises for andre (Moyaho et al., 2017; Palagi et al., 2019). Hos menneskelige og ikke-menneskelige primater, avhengig av art, når gjesping blir vist for andre, kan det kommunisere trussel (Troisi et al., 1990; Deputte, 1994) og / eller fysiologiske og atferdsmessige endringer (Provine et al., 1987 ; Leone et al., 2015; Zannella et al., 2015). Gjenging er en sosialt modulert respons fordi den kan hemmes av faktisk – og ikke virtuell – sosial tilstedeværelse (Gallup et al., 2019) og fordi en gjesp kan utløses av andres gjesp, som et resultat av et kjent fenomen. som smittsom gjesping (Provine, 1989, 2005). Gape smitte kan fremkalles selv om gjesp høres men ikke sees (Arnott et al., 2009; Massen et al., 2015).
Hos mennesker er deres fylogenetisk nærmeste apeart (sjimpanser: Pan paniscus) ; bonobos: Pan troglodytes) og den afrikanske apen Theropithecus gelada, smittsom gjesping er ikke bare til stede (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; men se: Amici et al., 2014), men også sosialt modulert fordi gjespesvaret er høyest når visse kategorier av individer er involvert (f.eks. pårørende, gruppemedlemmer, dominerende; Palagi et al., 2009; Campbell og de Waal, 2011, 2014; Norscia og Palagi, 2011; Demuru og Palagi, 2012; Massen et al., 2012). To hovedargumenter har blitt presentert for å forklare denne sosiale asymmetrien i smittsom gjesping, som er gruppert i to hovedhypoteser: Emotional Bias Hypothesis (EBH), som knytter smittsom gjesping til emosjonell overføring, og Attentional Bias Hypothesis (ABH), som vurderer smittsom gjesp som en motorisk respons som er gjenstand for forskjeller i nedadgående oppmerksomhetsprosesser (Palagi et al., 2020).
EBH spår at den sosiale asymmetrien observert i gjesksmittehastigheter gjenspeiler forskjeller i de forskjellige sosial binding, en proxy for emosjonell binding, mellom individer. Denne hypotesen støttes av bevis for at gjesksmittefrekvenser følger en empatisk trend (sensu Preston og de Waal, 2002), og er høyest mellom individer som har et sterkest emosjonelt bånd. Spesielt fant Norscia og Palagi (2011) at smittefrekvenser hos mennesker er størst som svar på familie og venner enn som svar på bekjente og fremmede. Hos voksne sjimpanser er gjespesmitte høyere mellom i-gruppen sammenlignet med medlemmer utenfor gruppen (Campbell og de Waal, 2011) og i bonobos er gjeste smittefrekvensen størst mellom individer som tilknytter seg mer til hverandre (Demuru og Palagi, 2012). I en komparativ undersøkelse inkludert både mennesker og bonoboer, Palagi et al. (2014) fant at gjengesmittefrekvensen ble påvirket av forholdskvaliteten mellom individer mer enn av arten forsøkspersonene tilhørte. I tillegg øker gjesp smitte med alderen når evnen til å identifisere andres følelser øker og avtar med høy alder når slik evne synker (Wiggers og van Lieshout, 1985; Anderson og Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen og Anderson, 2011; Bartholomew og Cirulli, 2014). Gape smittehastigheter øker fra spedbarn til voksen alder også hos sjimpanser (Madsen og Persson, 2013).
ABH spår at den sosiale asymmetrien som er observert i gjespesmitte kan skyldes forskjeller i sosial, visuell oppmerksomhet (Massen og Gallup, 2017). Spesielt ville de høyeste nivåene av smittsom gjesp være på grunn av den ekstra, nedadgående, selektive visuelle oppmerksomheten til individer som er mer relevante for observatøren, for eksempel kjente fag, slik det forekommer hos mennesker og gelader, eller dominanter, som det forekommer i sjimpanser eller bonoboer (Yoon og Tennie, 2010; Massen et al., 2012; Massen og Gallup, 2017).I følge Massen og Gallup (2017) vil ABH støttes av eksisterende bevis på den forskjellige visuelle deteksjonen og den visuelle perceptive kodingen av ansikter til kjente og / eller gruppefag sammenlignet med ukjente (f.eks. Buttle og Raymond, 2003 ; Ganel og Goshen-Gottstein, 2004; Jackson og Raymond, 2006; Michel et al., 2006).
I denne studien analyserte vi data om gjesp samlet over 9 år på mennesker i deres naturlige omgivelser og vi ekstrapolerte tilfellene der gjespet som sendes av et motiv kan bli hørt, men ikke sett av en potensiell responder (auditiv gjesp). Ved å vurdere bare de tilfellene der den visuelle signaturen til gjespestimuleringen ikke var påviselig, verifiserte vi om den sosiale asymmetrien som tidligere ble observert i gjespesmitte, vedvarte eller ikke. Spesielt testet vi følgende alternative spådommer avledet fra de to hypotesene som er presentert ovenfor (EBH og ABH).
Prediksjon 1a: ifølge EBH påvirkes hastigheten på gjespesmitte av styrken til inter -individuelt sosialt bånd – en fullmakt til det følelsesmessige båndet – per se og ikke av en annen ovenfra og ned, selektiv visuell oppmerksomhet spesielt til bestemte individer. Hvis denne hypotesen støttes, forventer vi å observere den sosiale skjevheten også når den visuelle signaturen til gjespestimuleringen er ekskludert, og frekvensen av auditiv smittsom gjesping blir høyere mellom sterkt bundet sammenlignet med svakt bundet individer.
Prediksjon 1b: ifølge ABH vil de høyere nivåene av gjespesmitte mellom sterkt bundet sammenlignet med svakt bundet individer være knyttet til den nærmeste ovenfra og ned, selektiv visuell oppmerksomhet som individer betaler til individer som er relevante for dem, for eksempel familie og venner. Hvis denne hypotesen støttes, bør den sosiale skjevheten som observeres i gjespesmitte forsvinne når bare hørselsgjesp blir vurdert fordi den visuelle signaturen ikke kan følges av den potensielle responderen.
Materialer og metoder
Datainnsamling og operasjonelle definisjoner
For denne studien vurderte vi de vokaliserte gjespene som sendes ut av et emne som bare kunne høres – men ikke sees – av en potensiell responder (heretter: auditiv gjesp). Emitteren og de potensielle respondentene måtte være i et område på ≤5 m. Vokaliserte gjesp involverte bruk av vokalfold og gjespene som bare innebar tung inspirasjon / utløp ble ikke ansett som vokaliserte; 294 tilfeller av hørselsgjesp ble ekstrapolert fra et datasett på totalt 2001 gjesper som ble samlet over 9 år — fra 2010 til 2019 — ved å bruke prøvetakingsmetoden for alle forekomster (Altmann, 1974). Spesielt ble hørselsgjespene samlet fra november 2010 til mai 2019, fra 05.30 til 02.30, på humane kaukasiske forsøkspersoner, i alderen 18 til 77 år, under rutinemessige aktiviteter, for eksempel på arbeidsplasser, over måltider, under sosiale møter osv ., med fagene er uvitende om å bli observert og i fravær av noen åpenbar ekstern kilde til angst. Den auditive gjespedatabasen inkluderte 193 gyper-potensielle responderdyer. Avhengig av situasjonen ble informasjonen registrert, ubemerket, gjennom alfanumeriske koder og lagt inn direkte i beregningsark, skrevet i mobiltelefoner eller skrevet på papir, og deretter lagt inn i beregningsark for senere utarbeidelse. Grunnleggende informasjon som alder og forholdet mellom mennesker var kjent for forfatterne. De potensielle respondentene ble kodet som i tilstanden som ikke var synlig når hodet ble rotert 180 ° i forhold til avtrekkeren, eller når en fysisk, blokkerende hindring var tilstede som forhindret den potensielle responderen fra å se utløserens ansikt og kropp. Utløseren og responderen ble aldri helt isolert (for eksempel i to separate rom med lukkede dører) fra hverandre. Den sosiale nærheten ble samlet på fire nivåer: 0 = fremmede, som aldri hadde møtt før; 1 = bekjente, som utelukkende delte et indirekte forhold basert på et tredje eksternt element, det vil si arbeidsplikt (kolleger) eller venner til felles (venners venner); 2 = venner, ikke-relaterte individer som deler et direkte forhold, ikke bare relatert til et tredje eksternt element; 3 = faste partnere og pårørende (r ≥ 0,25). Tidligere litteratur rapporterer at gjespesvar kan fremkalles innen 5 minutter etter å ha sett andres gjesp (utløserens gjesp) (Provine, 1986), med et maksimum i første minutt (Provine, 2005; Palagi et al., 2014). Litteratur rapporterer også at fra det fjerde minutt er det størst sannsynlighet for autokorrelasjon (noe som betyr at tilstedeværelsen av en gjesp utført av et subjekt ved t0 øker sannsynligheten for å ha en annen gjesp av samme subjekt ved t (0 + X) hvor X er den økende tidsenheten; Kapitány og Nielsen, 2017). Derfor vurderte vi de gjespesvarene som skjedde innen et tidsvindu på 3 minutter fra gjespene som utløses av utløseren.For ytterligere å redusere autokorrelasjonsforstyrrelsen, i tilfelle en kjede av gjesp som avgis av utløseren (mer gjesp slippes ut i 3-minutters tidsvinduet) registrerte vi som svar bare den første gjesp som ble utført etter oppfatningen av den siste gjesp. Vi kodet en gjesp som «spontan» når ingen andre personer hadde gjespet i løpet av 5 minutter før gjespende hendelse.
Statistiske analyser
For analysene ble følgende variabler vurdert: forekomst av smitte, kodet som: 1 = tilstedeværelse, 0 = fravær; det sosiale båndet ble angitt med de fire nivåene definert ovenfor (0 = fremmede; 1 = bekjente; 2 = venner; 3 = pårørende); utløserens og observatørens kjønn ble merket som : M = mann, F = kvinne; aldersklassene til utløseren og responderen ble kodet som følger: yo = ungdom (18-24 år); annonse = voksen (25-64 år gammel); se = senior (over 65 år gammel) (Statistics Canada, 2009); tidslukerne ble kodet som følger: 1 = 05:30 – 09:00; 2 = 09:01 – 12:30; 3 = 12: 31–16: 00:00; 4 = 16: 01–19: 30 pm; 5 = 19: 31–23: 00 pm; 6 = 23: 01–02: 30 (Giganti og Zilli, 2011). Databasen (se tilleggsdatablad) inkluderte 84 hanner, 69 kvinner, 16 ungdommer (yo), 122 voksne (ad) og 15 eldre (se). For å teste om forekomsten av gjesp smitte ble påvirket av faktorene bånd (0 = fremmede; 1 = bekjente; 2 = venner; 3 = kin), utløserens kjønn (Trigger_sex), responderens kjønn (Responder_sex), utløserens alder (Trigger_ageclass) og responderens (Responder_ageclass) aldersklasse, og tidsluken (fra 1 til 6 ), brukte vi en generalisert lineær blandet modell (GLMM) som inkluderte disse fem prediktorene som faste effekter og utløsere (Trigger) og respondenter (Responder) identiteter som tilfeldige effekter. Vi monterte modellene i R (R Core Team, 2018; versjon 3.5.1) ved hjelp av funksjonen lmer til R-pakken lme4 (Bates et al., 2015). Vi etablerte betydningen av den fullstendige modellen i forhold til en nullmodell som bare inneholder tilfeldige effekter (Forstmeier og Schielzeth, 2011). Vi brukte en sannsynlighetsforholdstest (Dobson, 2002) for å teste denne signifikansen (ANOVA med argumentet «Chisq»). Vi beregnet p-verdiene for de enkelte prediktorene basert på sannsynlighetsforholdstester mellom full og null-modellen ved å bruke R -funksjon «drop1» (Barr et al., 2013). Ettersom svarvariabelen var binær, brukte vi en binomial feilfordeling. Vi testet om interaksjonen mellom kjønnene eller aldersklassene til utløseren og responderen var signifikant, men da de ikke var, inkluderte vi dem ikke i modellen. Vi brukte en flere kontrastpakker (multcomp) for å utføre alle parvise sammenligninger for hvert bindingsnivå med Tukey-testen (Bretz et al., 2010). Vi rapporterte de Bonferroni-justerte p-verdiene, estimat (Est), standardfeil (SE) og z-verdier.
Resultater

Figur 1. Linjediagram av effekten av det sosiale båndet mellom trigger og responder (X-akse) på gjennomsnittlig forekomst av akustisk gjespesmitte (Y-akse). Venner og pårørende viser signifikant høyere gjengesmittefrekvenser enn fremmede og bekjente (Tukey-test: venner versus fremmede p < 0.001; pårørende versus fremmede p < 0,001; venner kontra bekjente p < 0,001; pårørende versus bekjente p < 0,001; andre kombinasjoner, ns).
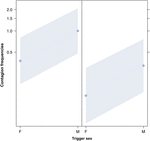
Figur 2. Linjeplott av effekten av utløserens kjønn (X-aksen) på akustisk gjespesmitte gjennomsnittlig forekomst (Y-aksen). Effekt av utløser-kjønn på akustisk gjespesmitte når responderen er en kvinne (høyre) og en mann (venstre). Hannes gjesp fremkaller flere gjesper enn hunnens uavhengig av kjønn av responderen (resultat av GLMM, p = 0,022). Poeng representerer effekten som responsvariabelen har på den uavhengige variabelen «utløserens kjønn,» basert på verdien som modellen forutsier. Fargede bånd viser 95% konfidensintervall.
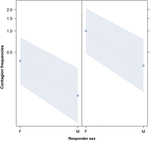
Figur 3. Linjediagram over effekten av kjønnet til responder (X-akse) på akustisk gjespesmitte gjennomsnittlig forekomst (Y-akse). Effekt av responderkjønn på akustisk gjespesmitte når utløseren er en kvinne (høyre) eller en mann (venstre). Kvinner reagerer betydelig mer enn menn uansett av utløserens kjønn (resultat av GLMM, p = 0,021). Poeng representerer effekten som responsvariabelen har på den uavhengige variabelen «kjønn av responderen», basert på verdien som modellen forutsier. Fargede bånd viser 95% konfidensintervall.
Diskusjon
Denne studien viser for første gang at gjesksmitte er betydelig påvirket av det sosiale båndet mellom individer (tabell 1) selv når de utløsende stimuli er auditiv gjesp, som vi definerte som vokaliserte gjesp som kunne høres, men ikke sees (visuell indikasjon, ikke påvisbar, auditiv signal påviselig). Spesielt er auditiv smittsom gjesping betydelig hyppigere mellom pårørende og venner enn mellom fremmede og bekjente (figur 1). Dette funnet støtter prediksjon 1a basert på EBH og ikke prediksjon 1b basert på ABH, noe som fører til konklusjonen at selektiv visuell oppmerksomhet ikke kan være hoveddriveren for sosial asymmetri observert i gjesksmittefrekvenser hos mennesker ovenfra og ned (Norscia og Palagi , 2011; Norscia et al., 2016a). Også utløserens kjønn og mottakerens kjønn hadde en signifikant effekt på gjespeinfeksjonsfrekvensen, med menn – som utløsere – som ble svart på av oftere enn kvinner (figur 2) og kvinner som svarte oftere på andres gjesp enn menn (figur 3).
I motsetning til Bartholomew og Cirulli (2014) fant vi ingen alderseffekt på gjespesmitte, mest sannsynlig fordi databasen vår om hørselsgjesp hadde en sterk forekomst av voksne (25–64 år). gammel). De høyeste nivåene av hørselsgjeving hos kvinner sammenlignet med menn bekrefter kjønnsskjevhet observert i naturalistiske forhold hos mennesker som er utsatt for gjesksmitte av Norscia et al. (2016a, b) når man vurderer et større datasett som også inkluderte gjesp som kunne sees av den potensielle responderen (med gjespende sensorisk modalitet – syn, hørsel eller begge deler – som ikke påvirker responsen). Kjønnsskjevheten stemmer også delvis overens med tidligere resultater oppnådd i kontrollerte innstillinger, inkludert den visuelle signalen (Chan og Tseng, 2017; men se Norscia og Palagi, 2011; Bartholomew og Cirulli, 2014). Det er blitt antatt at den høye graden av gjespesmitte hos kvinner kan informere emosjonell smitte (Norscia et al., 2016a), i lys av rapporterte høyere empatiske evner – relatert til barsel – av kvinner sammenlignet med menn (Christov-Moore et al. ., 2014). Imidlertid er dette problemet fortsatt under debatt fordi kulturelle forskjeller på tvers av menneskelige samfunn kan forme sosial bindingsdynamikk på en annen måte. Det er derfor komplisert, på dette stadiet av kunnskap, å løsne kulturelle faktorer, inter-personlig forholdskvalitet og kjønnsinnflytelse i fordelingen av gjespesmitte. Resultatene våre viser også at menn presterer bedre enn kvinner som utløsere, og den mest parsimonious hypotesen for dette kan være at menns vokalisering kan høres bedre i naturlige omgivelser, ofte preget av bakgrunnsstøy. Oppfatningen av stemmekjønn avhenger faktisk først og fremst av den grunnleggende frekvensen som i gjennomsnitt er lavere med en oktav hos mannlige stemmer enn kvinnestemmer, med lavere frekvensvokaliseringer som går lenger enn høyfrekvente (Marten og Marler, 1977; Latinus og Taylor, 2012) . Imidlertid, så vidt vi vet, er det ingen spesifikk studie som adresserer den mulige kjønnsforstyrrelsen i gjesphørbarhet, og videre undersøkelse med eksperimentelle studier i kontrollert tilstand er derfor nødvendig for å verifisere denne spekulasjonen.
I denne studien fant vi også ut at forskjellene i gjespesmitthet på tvers av kategorier (familie og venner, fremmede og bekjente) ikke kan forklares med forskjeller i ovenfra og ned, selektiv visuell oppmerksomhet. Dette funnet er i tråd med tidligere litteratur. Smittsom gjesping ser ut til å involvere hjerneområder som er mer relatert til det orienterende bunnen opp-nettverket enn de ovenfra og ned relaterte områdene. Videre er gjespesmitte hverken følsom overfor sensoriske signaler som er tilstede i signalet (auditive, visuelle eller audiovisuelle) (Arnott et al., 2009; Norscia og Palagi, 2011) eller påvirkes av det visuelle perspektivet til den utløsende stimulansen ( gjesp i retning 90 °, 180 ° og 270 ° er i stand til å utløse gjespingsresponser som frontal, 0 ° gjesp; Provine, 1989, 1996). Chan og Tseng (2017) fant at evnen til å oppdage et gjesp som sådan (perseptuell deteksjonsfølsomhet) var relatert til varigheten av blikket til øynene til stimulus som frigjorde ansiktet, men øye-blikkmønstre var ikke i stand til å modulere smittsom gjesp. I sjimpanser var smittsomme gjengefrekvenser høyest mellom samme gruppe enn forskjellige grupper, selv om respondentene så lenger på sjimpansevideoer utenfor gruppen (Campbell og de Waal, 2011). Argumentet om at visuell selektiv oppmerksomhet kan skjevgjøre smittehastigheter i en bestemt retning (personer som reagerer mer på familie enn fremmede) undergraves også av fraværet av noe spesifikt mønster for sosial oppmerksomhet hos menneskelige og ikke-menneskelige primater. Via øyesporing (brukt for å måle visningstiden) og ved å vise ukjente ansikter til deres eksperimentelle fag, Méary et al.(2014) observerte at mennesker var skjev mot ansikter fra egen rase, mens rhesus macaques oppmerksomhet ble mer tiltrukket av nye enn av samme artsansikter. Kawakami et al. (2014) observerte at mennesker tok mer hensyn til øynene til etniske medlemmer i gruppen og til nese og munn hos etniske medlemmer utenfor gruppen. Den samme studien avslørte også at visuell oppmerksomhet ikke var avhengig av målløpet. Ved å måle hvor lenge forsøkspersonene så på skjermen, Whitehouse et al. (2016) observerte at Barbary-makaker la mer vekt på å skrape videoer av ikke-fremmede enn fremmede individer, men bemerket også at innen ikke-fremmede, tok makakker mest oppmerksomhet til de individene som de delte en svak sosial binding med. Ved å måle blikkfrekvenser observerte Schino og Sciarretta (2016) at mandriller så mer på sine egne pårørende enn på ikke-pårørende, men også mer på dominerende enn på underordnede gruppekamerater. Derfor beskriver disse studiene (som brukes til å støtte ABH) ikke et enkelt mønster av selektiv oppmerksomhet. Et ytterligere viktig poeng å vurdere er selve definisjonen av fortrolighet og gruppemedlemskap som ble vedtatt av de fleste studiene som ble brukt for å støtte ABH (Massen og Gallup, 2017). Disse studiene viste bedre visuell gjenkjenning og visuell perseptiv koding av ansikter til kjente / i gruppefag sammenlignet med ukjente, men definerte fortrolighet og gruppemedlemskap ikke på grunnlag av de personlige forholdene mellom individer. I stedet ble fortrolighet eller gruppemedlemskap definert på grunnlag av indirekte kunnskap (f.eks. Foto av kjente personer eller et emne som allerede er vist i fasen før rettssaken) eller felles rase (f.eks. Buttle og Raymond, 2003; Ganel og Goshen- Gottstein, 2004; Jackson og Raymond, 2006; Michel et al., 2006). Denne definisjonen er greit for formålet med disse studiene, men det er ikke like bra hvis resultatene brukes til å foreslå alternative forklaringer på den påvirkningen som ekte sosial binding – basert på reelle forhold – kan ha på et fenomen, i dette tilfellet gjespe smitte . For eksempel Michel et al. (2006) observerte at kaukasiske og asiatiske personer bedre kunne gjenkjenne ansikter av samme rase, men denne forskjellen var ikke til stede hos asiatiske personer som hadde bodd i omtrent et år blant kaukasiere. Et annet diskusjonspunkt gjelder tilstedeværelsen av gjespesmitte hos barn med autismespektrumforstyrrelse (ASD), som ofte viser endringer i visuell oppmerksomhet (Richard og Lajiness-ONeill, 2015). Hos ASD-barn kan gjesksmitte være fraværende (Senju et al., 2007), svekket (Helt et al., 2010), eller lignende til barn som vanligvis utvikler seg når forsøkspersonene blir indusert til å omdirigere oppmerksomheten deres til videostimulus under eksperimentelle studier. (Usui et al., 2013). I en nylig studie har Mariscal et al. (2019) fant at gjespsmitte hos ASD-barn var positivt relatert til blodkonsentrasjonen av oksytocin, hormonet som er involvert i foreldrenes og sosiale tilknytning (Decety et al., 2016) og antydet at gjesksmitte hos ASD-barn kan være relatert til variabelt gjennomsnitt oksytocinkonsentrasjoner på tvers av forskjellige studiekohorter (Mariscal et al., 2019). Dette funnet er i tråd med EBH-hypotesen som knytter gjengesmittehastigheter til sosial binding, noe som kan gjenspeile emosjonell binding.
Konklusjon
Studien vår legger til diskusjonen om mekanismene som ligger til grunn for sosial asymmetri i gjespesmitte (for kritiske anmeldelser: se Adriaense et al., 2020; Palagi et al., 2020), ved å vise at gjesksmitte sannsynligvis er assosiert med nedenfra og opp, snarere enn med ovenfra og ned, selektiv oppmerksomhet. Bunn-opp-oppmerksomhet ledes først og fremst av den sensoriske oppfatningen av den fremkallende stimulansen mens ovenfra og ned, selektiv oppmerksomhet er en frivillig, vedvarende prosess der et bestemt element blir valgt internt og fokusert på eller undersøkt (Katsuki og Constantinidis, 2014). I denne forbindelse ble den akustiske stimulansen (auditiv gjesp) som ble avgitt av utløseren hørt, og kunne fremkalle et gjespende svar i mottakeren, selv om mottakeren ikke fulgte frivillig visuell oppmerksomhet til avtrekkeren. Videre var gjespingsfrekvensen sosialt modulert, med hørselsgavsmitte som høyest hos individer som var sterkest bundet til hverandre. Derfor er ikke selektiv oppmerksomhet ovenfra og ned ikke den viktigste driveren til den sosiale asymmetrien som er observert i gjespesmitte, noe som ser ut til å være et stimuleringsdrevet fenomen relatert til oppmerksomhetsprosesser nedenfra og opp. Videre undersøkelse er nødvendig for å forstå om og på hvilken måte andre former for oppmerksomhet eller pre-oppmerksomme stadier er i stand til å påvirke gjespesmitte.
Datatilgjengelighetserklæring
Datasettet som ble brukt for denne studien. er gitt i Supplementary Material.
Etikkerklæring
Studien som involverte menneskelige deltakere ble gjennomgått og godkjent av Comitato di Bioetica dAteneo (Universitetets bioetiske komité) —University of Turin (ref. nr. 451945).Skriftlig informert samtykke for deltakelse var ikke nødvendig for denne studien i samsvar med nasjonal lovgivning og institusjonelle krav.
Forfatterbidrag
IN og EP utførte datainnsamling og ble unnfanget og skrev manuskript. AZ hjalp til med datainnsamling og manuskriptrevisjon, figurer og tabeller. MG utførte statistiske analyser og skrev den relaterte delen av manuskriptet.
Interessekonflikt
Forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kunne tolkes som en potensiell interessekonflikt.
Anerkjennelser
Forfatterne ønsker å takke prof. Roberto Barbuti og prof. Cristina Giacoma for å ha støttet forskningen ved universitetet i Pisa og Universitetet i Torino, henholdsvis.
Supplerende materiale
Altmann, J. (1974). Observasjonsstudie av atferd: prøvetakingsmetoder. Atferd 49, 227–266. doi: 10.1163 / 156853974×00534
PubMed Abstract | CrossRef Fulltekst | Google Scholar
Anderson, J. R. og Meno, P. (2003). Psykologiske påvirkninger på gjesp hos barn. Curr. Psykol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J. og Cirulli, E. T. (2014). Individuell variasjon i smittsom gjesningsfølsomhet er svært stabil og i stor grad uforklarlig av empati eller andre kjente faktorer. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | CrossRef Fulltekst | Google Scholar
Bretz, F., Hothorn, T. og Westfall, P. (2010). Flere sammenligninger ved bruk av R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Etologisk studie av gjesping hos primater. I. Kvantitativ analyse og studie av årsakssammenheng hos to arter av gamleverdsaper (Cercocebus albigena og Macaca fascicularis). Etologi 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Full Text | Google Scholar
Dobson, A. J. (2002). En introduksjon til generaliserte lineære modeller, 2. utg. Boca Raton, FL: Chapman og Hall.
Google Scholar
Katsuki, F. og Constantinidis, C . (2014). Bunn-opp og ned-oppmerksomhet: forskjellige prosesser og overlappende nevrale systemer. Nevrolog 20, 509–521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef Fulltekst | Google Scholar
Singer, T. (2006). Nevronbasis og ontogeni av empati og tankelesing: gjennomgang av litteratur og implikasjoner for fremtidig forskning. Neurosci. Biobehav. Rev. 30, 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | CrossRef Fulltekst | Google Scholar
Statistics Canada (2009). Aldersgrupper, livssyklusgrupperinger. Ottawa: Statistics Canada.
Google Scholar