Bevezetés
Az ásítás a száj nyitásának, mély inspirációjának, rövid apnoéjának és többé-kevésbé lassú lejáratásának önkéntelen szekvenciája (Baenninger, 1997; Walusinski és Deputte, 2004; Guggisberg és mtsai, 2010; Krestel és mtsai, 2018). Kihíváskor az ásítást nem lehet teljesen elnyomni. Ezért sztereotip vagy reflexszerű mintaként határozták meg (Lehmann, 1979; Provine, 1986). A Homo sapiens-ben számos hipotézis változó támogatással került elő a spontán ásítás mechanizmusainak és funkcióinak magyarázatára, mint például oxigenizáció (hipoxia okozta légzési funkció), stresszel kapcsolatos viselkedés (izgalom okozta) vagy hőszabályozás (hipertermia okozta; Guggisberg és mtsai, 2010; Massen és mtsai, 2014; Gallup és Gallup, 2019). Fiziológiai válaszként az ásítást befolyásolhatják belső és külső tényezők, például a napszak (Giganti és Zilli, 2011) vagy koponyaűri / agyi hőmérséklet (Gallup és Eldakar, 2013).
Az ásítás képes legyen önirányított és / vagy mások számára megjelenített (Moyaho et al., 2017; Palagi et al., 2019). Emberi főemlősökben és főemlősökben, a fajtól függően, ha másoknak megmutatják az ásítást, kommunikálhat fenyegetéssel (Troisi et al., 1990; Deputte, 1994) és / vagy fiziológiai és viselkedési változásokkal (Provine et al., 1987 ; Leone és mtsai, 2015; Zannella és mtsai, 2015). Az emberekben az ásítás társadalmilag modulált válasz, mivel a tényleges – és nem virtuális – társadalmi jelenlét gátolhatja (Gallup et al., 2019), és mert az ásítást valaki más ásítás válthatja ki, egy ismert jelenség eredményeként. fertőző ásításként (Provine, 1989, 2005). Az ásításfertőzés akkor is kiváltható, ha az ásítást hallják, de nem látják (Arnott et al., 2009; Massen et al., 2015).
Az embereknél filogenetikailag a legközelebbi majomfajuk (csimpánzok: Pan paniscus ; bonobos: Pan troglodytes) és az afrikai Theropithecus gelada majom, a fertőző ásítás nemcsak jelen van (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; de lásd: Amici et al., 2014), hanem társadalmilag modulált, mert az ásító válasz akkor a legmagasabb, ha bizonyos egyének kategóriái érintettek (pl. rokonok, csoporttagok, dominánsok; Palagi et al., 2009; Campbell és de Waal, 2011, 2014; Norscia és Palagi, 2011; Demuru és Palagi, 2012; Massen és mtsai, 2012). Két fő érvet terjesztettek elő a fertőző ásítás társadalmi aszimmetriájának magyarázatára, amelyeket két fő hipotézisbe csoportosítottak: az érzelmi elfogultság hipotézisét (EBH), amely a fertőző ásítást összekapcsolja az érzelmi transzferrel, és a Figyelmi elfogultság hipotézist (ABH), amely figyelembe veszi fertőző ásítás, mint motoros válasz, amely függ a felülről lefelé irányuló figyelmi folyamatok különbségeitől (Palagi et al., 2020).
Az EBH előrejelzése szerint az ásítás fertőzési arányában megfigyelt társadalmi aszimmetria tükrözi a különböző különbségeket társadalmi kötődés, az érzelmi kötődés proxyja az egyének között. Ezt a hipotézist alátámasztják azok a bizonyítékok, amelyek szerint az ásításos fertőzési arány empatikus tendenciát követ (sensu Preston és de Waal, 2002), és a legmagasabb érzelmi kötelékkel rendelkező egyének között van a legmagasabb. Konkrétan Norscia és Palagi (2011) azt találta, hogy az emberekben az ásítás fertőzöttségi aránya a rokonokra és a barátokra adott válaszként a legnagyobb, mint az ismerősökre és az ismeretlenekre. Felnőtt csimpánzokban az ásítás fertőzése magasabb a csoporton belül, mint a csoporton kívüli tagok között (Campbell és de Waal, 2011), a bonobókban pedig az ásítás fertőzöttségi aránya a legnagyobb mértékben egymással kapcsolatban álló egyének között van (Demuru és Palagi, 2012). Az embereket és a bonobókat egyaránt magában foglaló összehasonlító vizsgálatban Palagi és mtsai. (2014) megállapította, hogy az ásítás fertőzöttségi rátákat az egyének közötti kapcsolat minősége jobban befolyásolta, mint az a faj, amelyhez az alanyok tartoztak. Ezenkívül az embereknél az ásítás fertőzése az életkor előrehaladtával növekszik, amikor az idős kor előrehaladtával más érzelmek azonosításának képessége növekszik és csökken, amikor ez a képesség csökken (Wiggers és van Lieshout, 1985; Anderson és Meno, 2003; Saxe és mtsai, 2004; Singer, 2006; Millen és Anderson, 2011; Bartholomew és Cirulli, 2014). Az ásítás fertőzöttségi aránya csecsemőktől felnőttkorig nő a csimpánzoknál is (Madsen és Persson, 2013).
Az ABH előrejelzése szerint az ásítás fertőzésében megfigyelt társadalmi aszimmetria oka lehet a társadalmi, vizuális figyelem (Massen és Gallup, 2017). Különösen a fertőző ásítás legmagasabb szintje annak az extra felülről lefelé irányuló, szelektív vizuális figyelemnek tudható be, amely a megfigyelő számára relevánsabb egyénekre irányul, például ismerős alanyokra, mivel ez előfordul az embereknél és a geladáknál, vagy a dominánsoknál. csimpánzokban vagy bonobókban fordul elő (Yoon és Tennie, 2010; Massen et al., 2012; Massen és Gallup, 2017).Massen és Gallup (2017) szerint az ABH-t a meglévő bizonyítékok támasztanák alá az ismerős és / vagy csoporton belüli alanyok arcainak eltérő vizuális észleléséről és vizuális percepciós kódolásáról az ismeretlenekhez képest (pl. Buttle és Raymond, 2003 ; Ganel és Goshen-Gottstein, 2004; Jackson és Raymond, 2006; Michel és mtsai, 2006).
Ebben a tanulmányban elemeztük az ásításról 9 év alatt gyűjtött adatokat az emberekről természetes környezetükben és extrapoláltuk azokat az eseteket, amikor az alany által kibocsátott ásítás hallható volt, de a potenciális válaszadó nem láthatta (hallási ásítás). Csak azoknak az eseteknek a figyelembevételével, amelyekben az ásító inger vizuális jelzése nem volt kimutatható, meggyőződtünk arról, hogy fennmaradt-e a társadalmi aszimmetria, amelyet korábban megfigyeltünk az ásítás fertőzési arányában. Különösen az alábbi alternatív jóslatokat teszteltük, amelyek a fentebb bemutatott két hipotézisből származnak (EBH és ABH).
1a. Jóslat: az EBH szerint az ásítás fertőzés mértékét befolyásolja az -egyéni társadalmi kötelék – az érzelmi kötelék proxyja – önmagában, és nem különösebb felülről lefelé irányuló, szelektív vizuális figyelem, amelyet bizonyos személyekre fordítanak. Ha ezt a hipotézist alátámasztjuk, akkor várhatóan megfigyeljük a társadalmi elfogultságot akkor is, ha az ásító inger vizuális jelét kizárjuk, és a hallási fertőző ásítás aránya magasabb lesz az erősen kötődő és a gyengén kötődő egyének között.
1b. Jóslat: az ABH szerint az erősen kötődő ásításos fertőzés magasabb szintje a gyengén kötődő egyénekhez képest a legközelebbi felülről lefelé irányuló, szelektív vizuális figyelemhez kapcsolódik, amelyet az egyének odafigyelnek a számukra releváns egyénekre, pl. barátok. Ha ezt a hipotézist alátámasztják, akkor az ásításfertőzésben megfigyelt társadalmi elfogultságnak el kell tűnnie, ha csak a hallásbeli ásításokat vesszük figyelembe, mivel a vizuális jelzésre a potenciális válaszadó nem képes figyelni.
Anyagok és módszerek
Adatgyűjtés és működési meghatározások
Ehhez a tanulmányhoz olyan alany által kibocsátott hangos ásításokat vettünk figyelembe, amelyet csak a potenciális válaszadó hallhatott meg – de nem láthatott – (a továbbiakban: hallási ásítások). A sugárzónak és a potenciális válaszadóknak ≤5 m tartományban kellett lenniük. A vokalizált ásítások vokális redők használatával jártak, és az ásításokat, amelyek csak súlyos inspirációt / kilégzést jelentettek, nem tekintettünk vokalizáltnak; 294 hallási ásítást extrapoláltak a 2001-es ásítások összesen 9 év alatt – 2010-től 2019-ig – összegyűjtött adatkészletéből az összes előfordulás mintavételi módszer alkalmazásával (Altmann, 1974). Konkrétan, hallási ásításokat gyűjtöttek 2010 novemberétől 2019 májusáig, 05.30 és 02.30 között emberi kaukázusi alanyokról, 18 és 77 év közötti, rutinszerű tevékenységeik során, például munkahelyen, étkezés közben, társadalmi találkozókon stb. ., az alanyok nincsenek tudatában megfigyelésüknek és nyilvánvaló külső szorongási források hiányában. Az auditív ásítás adatbázis 193 ásító-potenciális válaszadó diadát tartalmazott. A helyzet függvényében az információkat alfanumerikus kódokon keresztül rögzítették, nem vették figyelembe, és közvetlenül számítási lapokba vitték be, gépelték mobiltelefonra vagy papírra, majd későbbi kidolgozás céljából számítási lapokba vitték őket. Az alapinformációk, mint például az életkor és az emberek közötti kapcsolat, ismertek voltak a szerzők előtt. A potenciális válaszadókat úgy kódolták, mint a nem látás állapotát, amikor a fejüket 180 ° -kal elforgatták a ravasztól, vagy amikor fizikai, látást blokkoló akadály volt jelen, amely megakadályozta, hogy a potenciális válaszadó meglássa a ravasz arcát és testét. A kiváltó és a válaszadó soha nem volt teljesen elszigetelve (például két külön zárt ajtós helyiségben) egymástól. A társadalmi közelség négy szinten gyűlt össze: 0 = idegenek, akik még soha nem találkoztak; 1 = olyan ismerősök, akiknek csak közvetett kapcsolata volt egy harmadik külső elem alapján, vagyis a munkaköri kötelesség (kollégák) vagy közös barátok (barátok barátai); 2 = barátok, nem rokon személyek, akik közvetlen kapcsolatban állnak, és nem kizárólag egy harmadik külső elemhez kapcsolódnak; 3 = rendszeres partnerek és rokonok (r ≥ 0,25). Korábbi szakirodalom arról számolt be, hogy az ásító válaszok 5 percen belül kiválthatók, miután megnézték valaki más ásítását (a ravasz ásítását) (Provine, 1986), az első percben maximálisan (Provine, 2005; Palagi et al., 2014). Az irodalom arról is beszámol, hogy a negyedik perctől kezdve az autokorreláció valószínűsége a legnagyobb (ez azt jelenti, hogy egy alany által t0-nál végrehajtott ásítás jelenléte növeli annak valószínűségét, hogy ugyanaz a szubjektum t (0 + X) -nél újabb ásítást végezzen, ahol X az egyre növekvő időegység; Kapitány és Nielsen, 2017). Ezért figyelembe vettük az ásító válaszokat, amelyek 3 percen belül bekövetkeztek a trigger által kibocsátott ásítástól.Az autokorrelációs torzítás további csökkentése érdekében a ravasz által kibocsátott ásításlánc (több ásítás adódik a 3 perces időablakban) válaszként csak az utolsó ásítás észlelése után végrehajtott első ásítást regisztráltuk. Az ásítást “spontánnak” kódoltuk, amikor az ásítás eseményt megelőző 5 percben egyetlen másik személy sem ásított.
Statisztikai elemzések
Az elemzésekhez a következő változókat vettük figyelembe: előfordulás fertőzés, kódolva: 1 = jelenlét, 0 = hiány; a társadalmi köteléket a fent meghatározott négy szinttel vezették be (0 = idegenek; 1 = ismerősök; 2 = barátok; 3 = rokonok); a kiváltó és a megfigyelő nemét : M = férfi, F = nő; a kiváltó és a válaszadó korosztályait a következőképpen kódolták: yo = fiatal (18–24 éves); ad = felnőtt (25–64 éves); se = idősebb (fent 65 évesek) (Canada Statistics, 2009); az időréseket a következőképpen kódolták: 1 = 05: 30–09: 00; 2 = 09:01–12: 30; 3 = 12: 31–16: 00:00; 4 = 16: 01–19: 30; 5 = 19: 31–23: 00 pm; 6 = 23: 01–02: 30 (Giganti és Zilli, 2011). Az adatbázis (lásd Kiegészítő adatlap 84 férfi, 69 nő, 16 fiatal (yo), 122 felnőtt (ad) és 15 senior (se). A fertőzést a kötődési tényezők befolyásolták (0 = idegenek; 1 = ismerősök; 2 = barátok; 3 = rokonság), a ravasz neme (Trigger_sex), a válaszadó neme (Responder_sex), a trigger korosztálya (Trigger_ageclass) és a válaszadó korosztálya (Responder_ageclass), valamint az időrés (1-től 6-ig) ), egy általánosított lineáris vegyes modellt (GLMM) használtunk, amely ezt az öt előrejelzőt rögzített hatásként, kiváltó tényezőkként (Trigger) és válaszadók (Responder) identitásként véletlenszerű hatásokként foglalta magában. A modelleket R-be illesztettük (R Core Team, 2018; 3.5.1 verzió) az R-csomag lme4 funkciójának felhasználásával (Bates et al., 2015). Megállapítottuk a teljes modell jelentőségét egy csak véletlenszerű hatásokat tartalmazó null modellel összehasonlítva (Forstmeier és Schielzeth, 2011). Ennek a szignifikanciának a teszteléséhez valószínűségi arányt tesztet alkalmaztunk (Dobson, 2002) (ANOVA “Chisq” argumentummal). Az R használatával kiszámítottuk az egyes prediktorok p-értékeit a teljes és a null modell közötti valószínűségi arány tesztek alapján. -funkció “drop1” (Barr et al., 2013). Mivel a válaszváltozó bináris volt, binomiális hibaeloszlást használtunk. Kipróbáltuk, hogy a kiváltó tényező és a válaszadó nemek, illetve korosztályai közötti kölcsönhatás jelentős volt-e, de mivel nem voltak, nem vettük fel őket a modellbe. Többszörös kontrasztcsomagot (multcomp) használtunk az összes kötési szint összes páros összehasonlítására a Tukey-próbával (Bretz et al., 2010). Beszámoltunk a Bonferroni-korrigált p-értékekről, becslésről (Est), standard hibáról (SE) és z-értékekről.
Eredmények

1. ábra Vonalrajz a kiváltó és a válaszadó (X tengely) közötti társadalmi kötelék hatásának az akusztikus ásításfertőzés átlagos előfordulására (Y tengely). A barátok és rokonok szignifikánsan magasabb ásításos fertőzési gyakoriságot mutatnak, mint az idegenek és ismerősök (Tukey-teszt: barátok az idegenekkel szemben p < 0,001; rokonok az idegenekkel szemben p < 0,001; barátok kontra ismerősök p < 0,001; rokonok szemben ismerősök p < 0,001; egyéb kombinációk, ns).
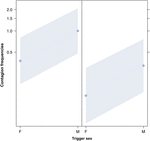
2. ábra a ravasz (X tengely) nemének hatása az akusztikus ásítás fertőzés átlagos előfordulására (Y tengely). A kiváltó nem hatása az akusztikus ásítás fertőzésre, amikor a válaszadó nő (jobb) és férfi (bal). A hím ásítások több ásítást váltanak ki, mint a nők, a válaszadó nemétől függetlenül (a GLMM eredménye, p = 0,022). A pontok azt a hatást mutatják, amelyet a válaszváltozó gyakorol a független változó “kiváltó nemére” a modell által megjósolt érték alapján. A színes sávok a 95% -os megbízhatósági intervallumot mutatják.
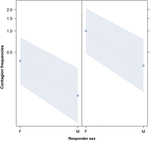
3. ábra Vonalrajz a válaszadó (X-tengely) az akusztikus ásítás fertőzés átlagos előfordulásán (Y-tengely). A válaszadó nem hatása az akusztikus ásítás-fertőzésre, amikor a kiváltó ok nő (jobb) vagy hím (bal). A nőstények lényegesen jobban reagálnak, mint a hímek A pontok azt a hatást mutatják, amelyet a válaszváltozó gyakorol a független “válaszadó nemére”, a modell által megjósolt érték alapján. A színes sávok a 95% -os konfidencia intervallumot mutatják.
Megbeszélés
Ez a tanulmány először mutatja be, hogy az ásító fertőzést jelentősen befolyásolja a társadalmi kötelék az egyének között (1. táblázat), még akkor is, ha a kiváltó ingerek hallási ásítások, amelyeket vokalizált ásításként definiáltunk, amelyek hallhatók, de nem láthatók (vizuális jel nem észlelhető, halló jel detektálható). Különösen a hallási fertőző ásítás lényegesen gyakoribb a rokonok és a barátok között, mint idegen és ismerős között (1. ábra). Ez a megállapítás alátámasztja az 1a előrejelzést az EBH alapján, és nem az 1b előrejelzést az ABH alapján, ami arra a következtetésre vezet, hogy az embereknél a felülről lefelé irányuló, szelektív vizuális figyelem nem lehet a fő hajtóereje az ásításfertőzési arányokban megfigyelt társadalmi aszimmetriának (Norscia és Palagi , 2011; Norscia et al., 2016a). A kiváltó ok és a vevő neme is jelentősen befolyásolta az ásítás fertőzöttségi arányát, a férfiakra – mint kiváltókra – mások reagálnak gyakrabban, mint a nők (2. ábra), a nők pedig gyakrabban reagálnak mások ásításaira, mint férfiak (3. ábra).
Bartholomew és Cirulli (2014) adatokkal ellentétben nem találtunk életkor-hatást az ásítás fertőzésre, valószínűleg azért, mert a hallási ásításokra vonatkozó adatbázisunkban a felnőttek (25–64 évesek) erősen elterjedtek. régi). A nőknél a legmagasabb szintű hallási ásításfertőzés megerősíti a nemi elfogultságot, amelyet Norscia és munkatársai naturalisztikus körülmények között figyeltek meg az ásítás fertőzésre fogékony embereken. (2016a, b) egy nagyobb adatkészlet megfontolásakor, amely magában foglalja a potenciális válaszadó számára látható ásításokat is (ásító érzékszervi modalitással – látás, hallás vagy mindkettő – nem befolyásolja a választ). A nemi elfogultság részben egyezik az ellenőrzött körülmények között elért korábbi eredményekkel, beleértve a vizuális jelzést is (Chan és Tseng, 2017; de lásd Norscia és Palagi, 2011; Bartholomew és Cirulli, 2014). Feltételezik, hogy a nők nagyfokú ásításfertőzése az érzelmi fertőzésre utalhat (Norscia et al., 2016a), a nők állítólag magasabb – az anyasághoz kapcsolódó – empatikus képességeinek fényében, mint a férfiak (Christov-Moore et al. ., 2014). Ez a kérdés azonban még mindig vita tárgya, mert az emberi társadalmak kulturális különbségei másképp formálhatják a társadalmi kötődési dinamikát. Ezért a tudásnak ebben a szakaszában bonyolult a kulturális tényezők, a személyek közötti kapcsolat minőségének és a nemek közötti befolyásolás szétválasztása az ásító fertőzés eloszlásában. Eredményeink azt is mutatják, hogy a férfiak kiváltóként jobban teljesítenek, mint a nők, és ennek a legszimpatikusabb hipotézise az lehet, hogy a férfiak hangja jobban hallható természetes körülmények között, gyakran háttérzajok jellemzik. Valójában a hangnem észlelése elsősorban arra az alapvető frekvenciára támaszkodik, amely átlagosan egy oktávval alacsonyabb a férfi, mint a női hangokban, és az alacsonyabb frekvenciájú hangzások tovább haladnak, mint a magas frekvenciájúak (Marten és Marler, 1977; Latinus és Taylor, 2012) . Tudomásunk szerint azonban nincs külön tanulmány, amely az ásítás hallhatóságának lehetséges nemi elfogultságával foglalkozna, ezért további ellenőrzések szükségesek kontrollált állapotú kísérleti kísérletekkel ennek a spekulációnak az igazolásához.
Ebben a tanulmányban azt is megtaláltuk hogy az ásításos fertőzöttség arányának különbségei a kategóriák (család és barátok, idegenek és ismerősök) között nem magyarázhatók a felülről lefelé irányuló, szelektív vizuális figyelem különbségeivel. Ez a megállapítás összhangban van a korábbi szakirodalommal. Úgy tűnik, hogy a fertőző ásítás olyan agyi területeket érint, amelyek jobban kapcsolódnak a tájoló-alulról felfelé irányuló hálózathoz, mint a felülről lefelé kapcsolódó területek. Sőt, az ásító fertőzés sem érzékeny a szignálban (hallási, vizuális vagy audiovizuális) jelen lévő szenzoros jelekre (Arnott és mtsai., 2009; Norscia és Palagi, 2011), és nem befolyásolja a kiváltó inger vizuális perspektívája ( a 90 °, 180 ° és 270 ° tájolású ásítások frontális, 0 ° -os ásításként képesek kiváltani az ásítás válaszait; Provine, 1989, 1996). Chan és Tseng (2017) megállapította, hogy az ásítás mint olyan észlelésének képessége (perceptuális detektálási érzékenység) összefüggésben állt az ingert felszabadító arc szemének tekintetével, de a szem-tekintet minták nem voltak képesek modulálni a fertőző ásítást. A csimpánzokban a fertőző ásítás gyakorisága a legnagyobb volt az azonos csoportok, mint a különböző csoportok egyének között, még akkor is, ha a válaszadók hosszabb ideig nézték a csoporton kívüli csimpánz videókat (Campbell és de Waal, 2011). Az az érv, miszerint a vizuális szelektív figyelem előrehaladhatja a fertőzés arányát egy adott irányban (az alanyok jobban reagálnak a családra, mint az idegenekre), aláássa az is, hogy az emberi főemlősökön és nem főemlősökön nincs semmiféle társadalmi figyelem. Szemkövetéssel (a nézési idő mérésére) és ismeretlen arcok megmutatásával kísérleti alanyaiknak, Méary et al.(2014) megfigyelte, hogy az emberek ferdén helyezkedtek el a saját fajuk arcai felé, míg a rhesus makákók figyelmét inkább az új vonzotta, mint az azonos fajú arcok. Kawakami és mtsai. (2014) megfigyelte, hogy az emberi alanyok nagyobb figyelmet fordítottak a csoporton belüli etnikai tagok szemére, valamint a csoporton kívüli tagok orrára és szájára. Ugyanez a tanulmány azt is feltárta, hogy a vizuális figyelem nem a célversenytől függ. Annak mérésével, hogy a kísérleti alanyok mennyi ideig nézték a képernyőt, Whitehouse et al. (2016) megfigyelte, hogy a barbár makákók nagyobb figyelmet fordítottak a nem idegen, mint az idegen személyek videóinak kaparására, de megjegyezték azt is, hogy a nem idegeneknél a makákók leginkább azokra az egyénekre figyeltek, akikkel gyenge társadalmi kötődésben voltak. A pillantási arányok mérésével Schino és Sciarretta (2016) megfigyelték, hogy a mandrillok inkább a saját rokonságukra tekintettek, mint a nem rokonokra, de inkább a dominánsra, mint az alárendelt csoporttársakra. Ezért ezek a tanulmányok (amelyeket az ABH alátámasztására használtak) egyetlen szelektív figyelemmintát sem írnak le. Egy további fontos szempont, amelyet figyelembe kell venni, az ABH támogatására használt tanulmányok többségében a családiasság és a csoporttagság meghatározása (Massen és Gallup, 2017). Ezek a tanulmányok jobb vizuális észlelést és az ismerős / csoporton belüli alanyok arcainak vizuális percepciós kódolását mutatták az ismeretlenekhez képest, de az ismeretséget és a csoporttagságot nem az egyének közötti személyes kapcsolatok alapján határozták meg. Ehelyett a családiasságot vagy a csoporttagságot közvetett ismeretek (pl. Híres emberek fényképe vagy egy tárgy, amelyet már a tárgyalás előtti szakaszban bemutattak) vagy közös faj (pl. Buttle és Raymond, 2003; Ganel és Goshen- Gottstein, 2004; Jackson és Raymond, 2006; Michel és mtsai, 2006). Ez a meghatározás e tanulmányok szempontjából rendben van, de nem annyira finom, ha az eredmények alapján alternatív magyarázatokat javasolunk arra a hatásra, amelyet a valós társadalmi kötődés – a valós kapcsolatokon alapulva – befolyásolhat egy jelenségre, ebben az esetben ásító fertőzésre . Például Michel és mtsai. (2006) megállapította, hogy a kaukázusi és az ázsiai alanyok jobban felismerik az azonos fajú arcokat, de ez a különbség nem volt jelen azoknál az ázsiai alanyoknál, akik körülbelül egy éve éltek a kaukázusiak körében. A vita egy másik pontja ásításfertőzés jelenlétét érinti az autista spektrumzavarral (ASD) szenvedő gyermekeknél, amelyek gyakran mutatják a vizuális figyelem megváltozását (Richard és Lajiness-O’Neill, 2015). ASD-s gyermekeknél az ásításfertőzés hiányozhat (Senju és mtsai, 2007), károsodhat (Helt és mtsai, 2010), vagy hasonló a tipikusan fejlődő gyermekekhez, amikor a kísérleti kísérletek során arra ösztönzik az alanyokat, hogy irányítsák figyelmüket a videoingerre (Usui és mtsai., 2013). Egy nemrégiben készült tanulmányban Mariscal et al. (2019) megállapította, hogy az ásító fertőzés ASD gyermekeknél pozitív kapcsolatban állt az oxitocin, a szülői és társadalmi kötődésben szerepet játszó hormon vérkoncentrációjával (Decety et al., 2016), és azt állította, hogy az ASD gyermekek ásításfertőződése összefüggésben lehet a változó átlaggal oxitocin koncentráció a különböző vizsgálati kohorszokban (Mariscal et al., 2019). Ez a megállapítás összhangban áll az EBH hipotézisével, amely összekapcsolja az ásítás terjedési arányát a társadalmi kötődéssel, amely tükrözi az érzelmi kötődést.
Következtetés
Vizsgálatunk kiegészíti a társadalmi aszimmetria az ásító fertőzésben (kritikai áttekintéshez: lásd Adriaense et al., 2020; Palagi et al., 2020), megmutatva, hogy az ásító fertőzés valószínűleg alulról felfelé, nem pedig felülről lefelé irányuló, szelektív figyelemmel társul. Az alulról felfelé irányuló figyelmet elsősorban a kiváltó inger érzékszervi észlelése vezeti, míg a felülről lefelé irányuló, szelektív figyelem önkéntes, tartós folyamat, amelyben egy adott elemet belül választanak ki, és összpontosítanak vagy megvizsgálnak (Katsuki és Constantinidis, 2014). Ebben a tekintetben a ravasz által kibocsátott akusztikus inger (hallási ásítás) hallható volt, és ásító választ váltott ki a vevőben, annak ellenére, hogy a vevő nem figyelt semmilyen önkéntes vizuális figyelmet a ravaszra. Ráadásul az ásítás válaszreakciói arányosan szociálisan moduláltak voltak, a hallási ásítás fertőzése azokban az egyénekben volt a legmagasabb, akik a legerősebben kapcsolódtak egymáshoz. Ennélfogva a felülről lefelé irányuló szelektív figyelem nem az ásító fertőzésben megfigyelt társadalmi aszimmetria fő mozgatórugója, amely úgy tűnik, hogy az alulról felfelé irányuló figyelem folyamataival kapcsolatos stimulus által vezérelt jelenség. További vizsgálatra van szükség annak megértéséhez, hogy a figyelem vagy az előzetes figyelmeztetés más formái képesek-e befolyásolni az ásításfertőzést és milyen módon.
Adatelérhetőségi nyilatkozat
A tanulmányhoz használt adatkészlet
Etikai nyilatkozat
Az emberi résztvevőket érintő tanulmányt a Comitato di Bioetica dAteneo (Egyetemi Bioetikai Bizottság) – Torinói Egyetem (ref. 451945).Ehhez a tanulmányhoz a nemzeti jogszabályokkal és az intézményi követelményekkel összhangban nem volt szükség írásos beleegyezésre a részvételhez.
Szerzői hozzászólások
Az IN és az EP elvégezte az adatgyűjtést, és megtervezte és megírta a tanulmányt. kézirat. Az AZ segített az adatgyűjtésben és a kéziratok felülvizsgálatában, az ábrákban és a táblázatokban. Az MG statisztikai elemzéseket végzett és megírta a kézirat kapcsolódó részét.
Összeférhetetlenség
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek lehetséges összeférhetetlenségként értelmezhető.
Köszönetnyilvánítás
A szerzők köszönetet mondanak Roberto Barbuti professzornak és Cristina Giacoma professzornak, hogy támogatták a Pisai Egyetemen folytatott kutatást és a Torinói Egyetem.
Kiegészítő anyagok
Altmann, J. (1974). A viselkedés megfigyeléses vizsgálata: mintavételi módszerek. Viselkedés 49, 227–266. doi: 10.1163 / 156853974×00534
PubMed Abstract | CrossRef teljes szöveg | Google Tudós
Anderson, J. R. és Meno, P. (2003). Pszichológiai hatások az ásításra gyermekeknél. Curr. Psychol. Lett. 11., 1–7.
Google Tudós
Bartholomew, A. J. és Cirulli, E. T. (2014). A fertőző ásítás fogékonyságának egyéni változásai nagyon stabilak, és empátia vagy más ismert tényezők nagyrészt megmagyarázhatatlanok. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | CrossRef teljes szöveg | Google Tudós
Bretz, F., Hothorn, T. és Westfall, P. (2010). Több összehasonlítás R. Boca Raton használatával, FL: Chapman & Hall.
Google Tudós
Deputte, BL (1994). A főemlősök ásításának etológiai vizsgálata. I. Az óvilági majmok két fajának (Cercocebus albigena és Macaca fascicularis) oksági mennyiségi elemzése és vizsgálata. Etológia 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef teljes szöveg | Google Tudós
Dobson, A. J. (2002). Bevezetés az általánosított lineáris modellekbe, 2. kiadás. Boca Raton, FL: Chapman és Hall.
Google Tudós
Katsuki, F. és Constantinidis, C. . (2014). Alulról felfelé és felülről lefelé irányuló figyelem: különböző folyamatok és átfedő idegrendszerek. Neuroscientist 20, 509–521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef teljes szöveg | Google Tudós
Singer, T. (2006). Az empátia és a gondolatolvasás neuronális alapjai és ontogenitása: az irodalom áttekintése és következményei a jövőbeni kutatásokra Neurosci. Biobehav. Jel. 30., 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
PubMed Abstract | CrossRef teljes szöveg | Google Tudós
Kanadai Statisztika (2009). Korosztályok, életciklus-csoportosítások. Ottawa: Kanadai statisztika.
Google Tudós