Introduzione
Lo sbadiglio è una sequenza involontaria di apertura della bocca, ispirazione profonda, breve apnea e espirazione più o meno lenta (Baenninger, 1997; Walusinski e Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Quando viene provocato, uno sbadiglio non può essere completamente soppresso. Pertanto, è stato definito come un modello stereotipato o simile a un riflesso (Lehmann, 1979; Provine, 1986). NellHomo sapiens sono state avanzate diverse ipotesi a supporto variabile per spiegare meccanismi e funzioni dello sbadiglio spontaneo, come lossigenazione (funzione respiratoria causata dallipossia), il comportamento correlato allo stress (causato dalleccitazione) o la termoregolazione (causata dallipertermia; Guggisberg et al., 2010; Massen et al., 2014; Gallup e Gallup, 2019). Essendo una risposta fisiologica, lo sbadiglio può essere influenzato da fattori interni ed esterni come lora del giorno (Giganti e Zilli, 2011) o la temperatura intracranica / cerebrale (Gallup e Eldakar, 2013).
Lo sbadiglio può essere auto-diretto e / o mostrato agli altri (Moyaho et al., 2017; Palagi et al., 2019). Nei primati umani e non umani, a seconda della specie, quando lo sbadiglio viene mostrato agli altri, può comunicare minaccia (Troisi et al., 1990; Deputte, 1994) e / o cambiamenti fisiologici e comportamentali (Provine et al., 1987 ; Leone et al., 2015; Zannella et al., 2015). Negli esseri umani, lo sbadiglio è una risposta socialmente modulata perché può essere inibito dalla presenza sociale effettiva e non virtuale (Gallup et al., 2019) e perché uno sbadiglio può essere innescato dallo sbadiglio di qualcun altro, come risultato di un fenomeno noto come sbadiglio contagioso (Provine, 1989, 2005). Il contagio dello sbadiglio può essere provocato anche se lo sbadiglio viene udito ma non visto (Arnott et al., 2009; Massen et al., 2015).
Negli esseri umani, le loro specie di scimmie filogeneticamente più vicine (scimpanzé: Pan paniscus ; bonobos: Pan troglodytes) e la scimmia africana Theropithecus gelada, lo sbadiglio contagioso non è solo presente (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; ma vedi: Amici et al., 2014) ma anche socialmente modulato perché la risposta allo sbadiglio è più alta quando sono coinvolte determinate categorie di individui (p. es., parenti, membri del gruppo, dominanti; Palagi et al., 2009; Campbell e de Waal, 2011, 2014; Norscia e Palagi, 2011; Demuru e Palagi, 2012; Massen et al., 2012). Due argomenti principali sono stati presentati per spiegare questa asimmetria sociale nello sbadiglio contagioso, che sono stati raggruppati in due ipotesi principali: lipotesi del pregiudizio emotivo (EBH), che collega lo sbadiglio contagioso al trasferimento emotivo, e lipotesi del pregiudizio attenzionale (ABH), che considera sbadiglio contagioso come risposta motoria soggetta a differenze nei processi attenzionali dallalto verso il basso (Palagi et al., 2020).
LEBH prevede che lasimmetria sociale osservata nei tassi di contagio di sbadiglio rifletta differenze nei diversi legame sociale, un proxy del legame emotivo, tra gli individui. Questa ipotesi è supportata dallevidenza che i tassi di contagio dello sbadiglio seguono una tendenza empatica (sensu Preston e de Waal, 2002), essendo più alti tra gli individui che condividono un legame emotivo più forte. In particolare, Norscia e Palagi (2011) hanno scoperto che negli esseri umani i tassi di contagio dello sbadiglio sono maggiori in risposta a parenti e amici che in risposta a conoscenti e sconosciuti. Negli scimpanzé adulti il contagio dello sbadiglio è maggiore tra i membri del gruppo rispetto a quelli esterni (Campbell e de Waal, 2011) e nei bonobo i tassi di contagio dello sbadiglio sono maggiori tra gli individui che si affiliano di più tra loro (Demuru e Palagi, 2012). In unindagine comparativa che includeva esseri umani e bonobo, Palagi et al. (2014) hanno scoperto che i tassi di contagio dello sbadiglio erano influenzati dalla qualità della relazione tra gli individui più che dalla specie a cui i soggetti appartenevano. Inoltre, negli esseri umani il contagio dello sbadiglio aumenta con letà quando la capacità di identificare le emozioni degli altri aumenta e diminuisce con la vecchiaia quando tale capacità diminuisce (Wiggers e van Lieshout, 1985; Anderson e Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen e Anderson, 2011; Bartholomew e Cirulli, 2014). I tassi di contagio dello sbadiglio aumentano dallinfanzia alletà adulta anche negli scimpanzé (Madsen e Persson, 2013).
LABH prevede che lasimmetria sociale osservata nel contagio dello sbadiglio possa essere dovuta a differenze nellattenzione sociale e visiva (Massen e Gallup, 2017). In particolare, i livelli più alti di sbadiglio contagioso sarebbero dovuti alla maggiore attenzione visiva selettiva dallalto verso il basso prestata agli individui che sono più rilevanti per losservatore, come i soggetti familiari, come si verifica negli esseri umani e nelle geladas, o si verifica negli scimpanzé o nei bonobo (Yoon e Tennie, 2010; Massen et al., 2012; Massen e Gallup, 2017).Secondo Massen e Gallup (2017), ABH sarebbe supportato dalle prove esistenti sulla diversa rilevazione visiva e codifica percettiva visiva di volti di soggetti familiari e / o in gruppo rispetto a quelli non familiari (ad esempio, Buttle e Raymond, 2003 ; Ganel e Goshen-Gottstein, 2004; Jackson e Raymond, 2006; Michel et al., 2006).
In questo studio, abbiamo analizzato i dati sullo sbadiglio raccolti in 9 anni sugli esseri umani nel loro ambiente naturale e abbiamo estrapolato i casi in cui lo sbadiglio emesso da un soggetto poteva essere udito ma non visto da un potenziale rispondente (sbadiglio uditivo). Considerando solo i casi in cui il segnale visivo dello stimolo di sbadiglio non era rilevabile, abbiamo verificato se lasimmetria sociale precedentemente osservata nei tassi di contagio di sbadiglio persistesse o meno. In particolare, abbiamo testato le seguenti previsioni alternative derivate dalle due ipotesi presentate sopra (EBH e ABH).
Predizione 1a: secondo lEBH, i tassi di contagio di sbadiglio sono influenzati dalla forza del – legame sociale individuale – un proxy del legame emotivo – di per sé e non da unattenzione visiva selettiva e dallalto verso il basso diversa prestata a determinati individui in particolare. Se questa ipotesi è supportata, ci aspettiamo di osservare il bias sociale anche quando il segnale visivo dello stimolo di sbadiglio è escluso e i tassi di sbadiglio contagioso uditivo saranno più alti tra individui fortemente legati rispetto a individui debolmente legati.
Predizione 1b: secondo lABH, i livelli più elevati di contagio di sbadiglio tra individui fortemente legati rispetto a soggetti debolmente legati sarebbero collegati alla più vicina attenzione visiva selettiva top-down che gli individui prestano agli individui che sono rilevanti per loro, ad esempio, la famiglia e amici. Se questa ipotesi è supportata, il bias sociale osservato nel contagio di sbadiglio dovrebbe scomparire quando si considerano solo gli sbadigli uditivi perché il segnale visivo non può essere seguito dal potenziale rispondente.
Materiali e metodi
Raccolta dei dati e definizioni operative
Per questo studio, abbiamo considerato gli sbadigli vocalizzati emessi da un soggetto che poteva essere solo ascoltato, ma non visto, da un potenziale risponditore (di seguito: sbadigli uditivi). Lemettitore e i potenziali risponditori dovevano trovarsi in un raggio di ≤5 m. Gli sbadigli vocalizzati prevedevano luso delle corde vocali e gli sbadigli che comportavano solo forti inspirazioni / espirazioni non erano considerati vocalizzati; 294 casi di sbadigli uditivi sono stati estrapolati da un set di dati di un totale di 2001 attacchi di sbadiglio raccolti in 9 anni, dal 2010 al 2019, utilizzando il metodo di campionamento di tutte le occorrenze (Altmann, 1974). Nello specifico, gli sbadigli uditivi sono stati raccolti da novembre 2010 a maggio 2019, dalle 05.30 alle 02.30, su soggetti umani caucasici, di età compresa tra 18 e 77 anni, durante le loro attività di routine, ad es. Nei luoghi di lavoro, durante i pasti, durante gli incontri sociali, ecc. ., con i soggetti inconsapevoli di essere osservati e in assenza di qualsiasi fonte esterna evidente di ansia. Il database dello sbadiglio uditivo includeva 193 diadi che rispondono con potenziale sbadiglio. A seconda della situazione, le informazioni sono state registrate, non annotate, tramite codici alfanumerici e inserite direttamente nei fogli di calcolo, digitate nei telefoni cellulari o scritte su carta, quindi inserite nei fogli di calcolo per la successiva elaborazione. Le informazioni di base come letà e il rapporto tra le persone erano note agli autori. I potenziali soccorritori sono stati codificati come nella condizione di non vista quando la loro testa era ruotata di 180 ° rispetto al grilletto o quando era presente un ostacolo fisico che bloccava la vista che impediva al potenziale rispondente di vedere il viso e il corpo del grilletto. Il grilletto e il risponditore non sono mai stati completamente isolati (ad esempio, in due stanze separate con porte chiuse) luno dallaltro. La vicinanza sociale è stata raccolta su quattro livelli: 0 = estranei, che non si erano mai incontrati prima; 1 = conoscenti, che condividevano esclusivamente un rapporto indiretto basato su un terzo elemento esterno, cioè il dovere di lavoro (colleghi) o amici in comune (amici di amici); 2 = amici, persone non imparentate che condividono un rapporto diretto non esclusivamente correlato a un terzo elemento esterno; 3 = partner regolari e parenti (r ≥ 0,25). La letteratura precedente riporta che le risposte di sbadiglio possono essere suscitate entro 5 minuti dopo aver visto lo sbadiglio di qualcun altro (lo sbadiglio del grilletto) (Provine, 1986), con un massimo nel primo minuto (Provine, 2005; Palagi et al., 2014). La letteratura riporta anche che dal quarto minuto cè una più alta probabilità di autocorrelazione (il che significa che la presenza di uno sbadiglio eseguito da un soggetto a t0 aumenta la probabilità di avere un altro sbadiglio dallo stesso soggetto at (0 + X) dove X è lunità di tempo crescente; Kapitány e Nielsen, 2017). Quindi, abbiamo considerato le risposte di sbadiglio che si verificano entro una finestra di tempo di 3 minuti dallo sbadiglio emesso dal trigger.Per ridurre ulteriormente il bias di autocorrelazione, in caso di catena di sbadigli emessi dal trigger (più sbadigli emessi nella finestra temporale di 3 min) abbiamo registrato come risposta solo il primo sbadiglio eseguito dopo la percezione dellultimo sbadiglio. Abbiamo codificato uno sbadiglio come “spontaneo” quando nessun altro soggetto aveva sbadigliato nei 5 minuti precedenti levento di sbadiglio.
Analisi statistiche
Per le analisi, sono state considerate le seguenti variabili: occorrenza di contagio, codificato come: 1 = presenza, 0 = assenza; il legame sociale è stato inserito con i quattro livelli sopra definiti (0 = estranei; 1 = conoscenti; 2 = amici; 3 = parenti); il sesso del trigger e dellosservatore sono stati etichettati come : M = maschio, F = femmina; le classi di età del trigger e del responder sono state codificate come segue: yo = giovani (18-24 anni); ad = adulto (25-64 anni); se = senior (sopra 65 anni) (Statistics Canada, 2009); le fasce orarie sono state codificate come segue: 1 = 05: 30–09: 00 am; 2 = 09:01 am-12: 30 pm; 3 = 12: 31–16: 00 pm; 4 = 16: 01–19: 30 pm; 5 = 19: 31–23: 00 pm; 6 = 23: 01–02: 30 (Giganti e Zilli, 2011). Il database (vedi Supplementary Data Sheet) inclusi 84 maschi, 69 femmine, 16 giovani (anni), 122 adulti (annuncio) e 15 anziani (se). Per verificare se il verificarsi di sbadigli il contagio è stato influenzato dai fattori legame (0 = estranei; 1 = conoscenti; 2 = amici; 3 = kin), sesso del trigger (Trigger_sex), sesso del rispondente (Responder_sex), classe di età del trigger (Trigger_ageclass) e fascia di età del risponditore (Responder_ageclass) e fascia oraria (da 1 a 6 ), abbiamo utilizzato un modello misto lineare generalizzato (GLMM) che includeva questi cinque predittori come effetti fissi e identità di trigger (Trigger) e responder (Responder) come effetti casuali. Abbiamo montato i modelli in R (R Core Team, 2018; versione 3.5.1) utilizzando la funzione lmer del pacchetto R lme4 (Bates et al., 2015). Abbiamo stabilito il significato del modello completo rispetto a un modello nullo che comprende solo gli effetti casuali (Forstmeier e Schielzeth, 2011). Abbiamo utilizzato un test del rapporto di verosimiglianza (Dobson, 2002) per verificare questa significatività (ANOVA con argomento “Chisq”). Abbiamo calcolato i valori p per i singoli predittori in base ai test del rapporto di verosimiglianza tra il modello completo e quello nullo utilizzando il -funzione “drop1” (Barr et al., 2013). Poiché la variabile di risposta era binaria, abbiamo utilizzato una distribuzione binomiale degli errori. Abbiamo verificato se linterazione tra i sessi o le classi di età del trigger e del responder fosse significativa, ma poiché non lo erano, non le abbiamo incluse nel modello. Abbiamo utilizzato un pacchetto di contrasto multiplo (multcomp) per eseguire tutti i confronti a coppie per ciascun livello di legame con il test di Tukey (Bretz et al., 2010). Abbiamo riportato i valori p aggiustati per Bonferroni, la stima (Est), lerrore standard (SE) e i valori z.
Risultati

Figura 1. Grafico a linee delleffetto del legame sociale tra trigger e responder (asse X) sulloccorrenza media del contagio di sbadiglio acustico (asse Y). Amici e parenti mostrano frequenze di contagio di sbadiglio significativamente più elevate rispetto a sconosciuti e conoscenti (test di Tukey: amici contro estranei p < 0,001; parenti contro estranei p < 0,001; amici contro conoscenti p < 0,001; parenti contro conoscenti p < 0,001; altre combinazioni, ns).
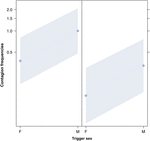
Figura 2. Grafico a linee di leffetto del sesso del trigger (asse X) sulloccorrenza media del contagio da sbadiglio acustico (asse Y). Effetto del sesso scatenante sul contagio da sbadiglio acustico quando il rispondente è una femmina (a destra) e un maschio (a sinistra). Gli sbadigli dei maschi provocano più sbadigli di quelli delle femmine indipendentemente dal sesso del rispondente (risultato del GLMM, p = 0,022). I punti rappresentano leffetto che la variabile di risposta ha sulla variabile indipendente “sesso dellattivatore”, in base al valore previsto dal modello. Le bande colorate mostrano lintervallo di confidenza del 95%.
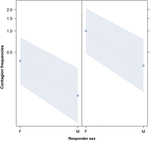
Figura 3. Grafico a linee delleffetto del sesso del risponditore (asse X) su insorgenza media di contagio da sbadiglio acustico (asse Y). Effetto del sesso del rispondente sul contagio da sbadiglio acustico quando il fattore scatenante è una femmina (destra) o un maschio (sinistra). Le femmine rispondono significativamente più dei maschi indipendentemente del sesso del trigger (risultato del GLMM, p = 0,021) I punti rappresentano leffetto che la variabile di risposta ha sulla variabile indipendente “sesso del rispondente”, in base al valore previsto dal modello. Le bande colorate mostrano lintervallo di confidenza del 95%.
Discussione
Questo studio mostra per la prima volta che il contagio dello sbadiglio è significativamente influenzato dal legame sociale tra gli individui (Tabella 1) anche quando gli stimoli scatenanti sono sbadigli uditivi, che abbiamo definito come sbadigli vocalizzati che potrebbero essere ascoltati ma non visti (segnale visivo non rilevabile, segnale uditivo rilevabile). In particolare, lo sbadiglio uditivo contagioso è significativamente più frequente tra parenti e amici che tra estranei e conoscenti (Figura 1). Questa scoperta supporta la previsione 1a basata sullEBH e non la predizione 1b basata sullABH, portando alla conclusione che negli esseri umani lattenzione visiva top-down, selettiva non può essere il principale motore dellasimmetria sociale osservata nei tassi di contagio di sbadiglio (Norscia e Palagi , 2011; Norscia et al., 2016a). Anche il sesso del soggetto scatenante e il sesso del ricevente hanno avuto un effetto significativo sui tassi di contagio di sbadiglio, con gli uomini – come fattori scatenanti – a cui hanno risposto gli altri più frequentemente delle donne (Figura 2) e le donne che rispondono più frequentemente agli sbadigli degli altri uomini (Figura 3).
Contrariamente a Bartholomew e Cirulli (2014), non abbiamo trovato alcun effetto delletà sul contagio dello sbadiglio, molto probabilmente perché il nostro database sugli sbadigli uditivi aveva una forte prevalenza di adulti (25-64 anni vecchio). I più alti livelli di contagio da sbadiglio uditivo nelle donne rispetto agli uomini confermano il bias di genere osservato in condizioni naturalistiche su esseri umani suscettibili al contagio di sbadiglio da Norscia et al. (2016a, b) quando si considera un set di dati più ampio che includeva anche sbadigli che potrebbero essere visti dal potenziale rispondente (con modalità sensoriali sbadiglianti – vista, udito o entrambi – che non influiscono sulla risposta). Il pregiudizio di genere è anche in parziale accordo con i risultati precedenti ottenuti in contesti controllati, incluso il segnale visivo (Chan e Tseng, 2017; ma vedi Norscia e Palagi, 2011; Bartholomew e Cirulli, 2014). È stato ipotizzato che lelevato grado di contagio dello sbadiglio nelle donne possa informare il contagio emotivo (Norscia et al., 2016a), alla luce delle capacità empatiche riferite più elevate, legate alla maternità, delle donne rispetto agli uomini (Christov-Moore et al. ., 2014). Tuttavia, questo problema è ancora in discussione perché le differenze culturali tra le società umane possono plasmare le dinamiche di legame sociale in un modo diverso. È quindi complicato, in questa fase della conoscenza, districare i fattori culturali, la qualità delle relazioni interpersonali e linfluenza di genere nella distribuzione del contagio di sbadiglio. I nostri risultati mostrano anche che gli uomini si comportano meglio delle donne come trigger e lipotesi più parsimoniosa potrebbe essere che le vocalizzazioni degli uomini possano essere ascoltate meglio in ambienti naturali, spesso caratterizzati da rumori di fondo. In effetti, la percezione del genere vocale si basa principalmente sulla frequenza fondamentale che è in media inferiore di unottava nelle voci maschili rispetto alle voci femminili, con vocalizzazioni a frequenza più bassa che viaggiano oltre quelle ad alta frequenza (Marten e Marler, 1977; Latinus e Taylor, 2012) . Tuttavia, a nostra conoscenza, non esiste uno studio specifico che affronti il possibile pregiudizio di genere nelludibilità dello sbadiglio e sono quindi necessarie ulteriori indagini con prove sperimentali in condizioni controllate per verificare questa speculazione.
In questo studio, abbiamo anche trovato che le differenze nei tassi di contagio dello sbadiglio tra le categorie (famiglia e amici, sconosciuti e conoscenti) non possono essere spiegate da differenze nellattenzione visiva selettiva dallalto verso il basso. Questo risultato è in linea con la letteratura precedente. Lo sbadiglio contagioso sembra coinvolgere le aree del cervello che sono più legate alla rete di orientamento dal basso verso lalto rispetto alle aree correlate dallalto verso il basso. Inoltre, il contagio dello sbadiglio non è né sensibile ai segnali sensoriali presenti nel segnale (uditivo, visivo o audiovisivo) (Arnott et al., 2009; Norscia e Palagi, 2011) né influenzato dalla prospettiva visiva dello stimolo scatenante ( sbadigli in orientamenti di 90 °, 180 ° e 270 ° sono in grado di innescare risposte di sbadiglio come sbadigli frontali, 0 °; Provine, 1989, 1996). Chan e Tseng (2017) hanno scoperto che la capacità di rilevare uno sbadiglio in quanto tale (sensibilità di rilevamento percettivo) era correlata alla durata dello sguardo verso gli occhi del viso che rilasciava lo stimolo, ma i modelli di sguardo-sguardo non erano in grado di modulare lo sbadiglio contagioso. Negli scimpanzé, le frequenze di sbadiglio contagiose erano più alte tra individui dello stesso gruppo rispetto a individui di gruppi diversi, anche se i soccorritori hanno guardato più a lungo i video di scimpanzé fuori gruppo (Campbell e de Waal, 2011). Largomento secondo cui lattenzione selettiva visiva può influenzare i tassi di contagio dello sbadiglio in una direzione specifica (soggetti che rispondono più alla famiglia che agli estranei) è anche minato dallassenza di qualsiasi modello specifico di attenzione sociale nei primati umani e non umani. Tramite eye-tracking (applicato per misurare il tempo di visione) e mostrando volti sconosciuti ai soggetti sperimentali, Méary et al.(2014) hanno osservato che gli umani erano distorti verso i volti della propria razza, mentre lattenzione dei macachi rhesus era più attratta dai volti nuovi che da quelli della stessa specie. Kawakami et al. (2014) hanno osservato che i soggetti umani prestavano maggiore attenzione agli occhi dei membri del gruppo etnico e al naso e alla bocca dei membri del gruppo etnico esterno. Lo stesso studio ha anche rivelato che lattenzione visiva non dipendeva dalla razza target. Misurando per quanto tempo i soggetti sperimentali hanno guardato lo schermo, Whitehouse et al. (2016) hanno osservato che i macachi barbareschi prestavano maggiore attenzione a graffiare video di individui non estranei rispetto a individui estranei, ma hanno anche notato che allinterno dei non estranei, i macachi prestavano maggiore attenzione a quegli individui con cui condividevano un debole legame sociale. Misurando i tassi di occhiata, Schino e Sciarretta (2016) hanno osservato che i mandrilli guardavano più ai propri parenti che ai non parenti, ma anche più ai compagni di gruppo dominante che a quelli subordinati. Pertanto, questi studi (utilizzati per supportare lABH) non descrivono un singolo modello di attenzione selettiva. Un altro punto importante da considerare è la definizione stessa di familiarità e appartenenza al gruppo adottata dalla maggior parte degli studi utilizzati per supportare ABH (Massen e Gallup, 2017). Questi studi hanno mostrato una migliore rilevazione visiva e codifica percettiva visiva di volti di soggetti familiari / in gruppo rispetto a quelli non familiari, ma hanno definito la familiarità e lappartenenza al gruppo non sulla base delle relazioni personali tra gli individui. Invece, la familiarità o lappartenenza a un gruppo sono state definite sulla base della conoscenza indiretta (ad esempio, foto di personaggi famosi o di un soggetto già mostrato nelle fasi preliminari) o di razza comune (ad esempio, Buttle e Raymond, 2003; Ganel e Goshen- Gottstein, 2004; Jackson e Raymond, 2006; Michel et al., 2006). Questa definizione va bene ai fini di questi studi, ma non è altrettanto valida se i risultati sono usati per proporre spiegazioni alternative per linfluenza che il legame sociale reale, basato su relazioni reali, può avere su un fenomeno, in questo caso sbadiglio contagio . Ad esempio, Michel et al. (2006) hanno osservato che i soggetti caucasici e asiatici potevano riconoscere meglio i volti della stessa razza, ma questa differenza non era presente nei soggetti asiatici che vivevano da circa un anno tra i caucasici. Un altro punto di discussione riguarda la presenza del contagio dello sbadiglio nei bambini con disturbo dello spettro autistico (ASD), che spesso mostrano alterazioni dellattenzione visiva (Richard e Lajiness-O’Neill, 2015). Nei bambini con ASD, il contagio dello sbadiglio può essere assente (Senju et al., 2007), alterato (Helt et al., 2010) o simile ai bambini in via di sviluppo tipico quando i soggetti sono indotti a reindirizzare la loro attenzione lo stimolo video durante le prove sperimentali (Usui et al., 2013). In uno studio recente, Mariscal et al. (2019) hanno scoperto che il contagio dello sbadiglio nei bambini con ASD era correlato positivamente alla concentrazione ematica di ossitocina, lormone coinvolto nellattaccamento genitoriale e sociale (Decety et al., 2016) e hanno ipotizzato che il contagio dello sbadiglio nei bambini con ASD possa essere correlato alla media variabile concentrazioni di ossitocina in diverse coorti di studio (Mariscal et al., 2019). Questa scoperta è in linea con lipotesi EBH che collega i tassi di contagio dello sbadiglio al legame sociale, che può riflettere il legame emotivo.
Conclusione
Il nostro studio si aggiunge alla discussione sui meccanismi alla base del asimmetria sociale nel contagio di sbadiglio (per una revisione critica: vedere Adriaense et al., 2020; Palagi et al., 2020), mostrando che il contagio di sbadiglio è probabilmente associato a unattenzione selettiva dal basso verso lalto, piuttosto che dallalto verso il basso. Lattenzione dal basso verso lalto è guidata principalmente dalla percezione sensoriale dello stimolo che suscita, mentre lattenzione selettiva dallalto verso il basso è un processo volontario e sostenuto in cui un particolare elemento viene selezionato internamente e focalizzato o esaminato (Katsuki e Constantinidis, 2014). A questo proposito, lo stimolo acustico (sbadiglio uditivo) emesso dal grilletto è stato sentito e potrebbe suscitare una risposta di sbadiglio nel ricevitore, anche se il ricevitore non stava prestando alcuna attenzione visiva volontaria al grilletto. Inoltre, i tassi di risposta allo sbadiglio erano socialmente modulati, con il contagio dello sbadiglio uditivo più alto negli individui che erano più fortemente legati luno allaltro. Quindi, lattenzione selettiva dallalto verso il basso non è il motore principale dellasimmetria sociale osservata nel contagio di sbadiglio, che sembra essere un fenomeno guidato dallo stimolo correlato ai processi di attenzione dal basso verso lalto. Sono necessarie ulteriori indagini per capire se e in che modo altre forme di attenzione o fasi di pre-attenzione sono in grado di influenzare il contagio dello sbadiglio.
Dichiarazione sulla disponibilità dei dati
Il set di dati utilizzato per questo studio è fornito nel Materiale Supplementare.
Dichiarazione Etica
Lo studio che ha coinvolto partecipanti umani è stato esaminato e approvato dal Comitato di Bioetica dAteneo (Comitato di Bioetica dellUniversità) – Università di Torino (rif . n. 451945).Il consenso informato scritto per la partecipazione non era richiesto per questo studio in conformità con la legislazione nazionale e i requisiti istituzionali.
Contributi dellautore
IN ed EP hanno effettuato la raccolta dei dati e concepito e scritto manoscritto. AZ ha aiutato con la raccolta dei dati e la revisione dei manoscritti, le figure e le tabelle. MG ha effettuato analisi statistiche e ha scritto la parte correlata del manoscritto.
Conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di rapporti commerciali o finanziari che potrebbero essere interpretato come un potenziale conflitto di interessi.
Ringraziamenti
Gli autori desiderano ringraziare il Prof. Roberto Barbuti e la Prof. Cristina Giacoma per aver sostenuto la ricerca presso lUniversità di Pisa e il Università di Torino, rispettivamente.
Supplementary Material
Altmann, J. (1974). Studio osservazionale del comportamento: metodi di campionamento. Comportamento 49, 227-266. doi: 10.1163 / 156853974×00534
Estratto di PubMed | Testo completo CrossRef | Google Scholar
Anderson, J. R. e Meno, P. (2003). Influenze psicologiche sullo sbadiglio nei bambini. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J. e Cirulli, E. T. (2014). La variazione individuale nella suscettibilità allo sbadiglio contagioso è altamente stabile e in gran parte inspiegabile dallempatia o da altri fattori noti. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
Estratto di PubMed | Testo completo CrossRef | Google Scholar
Bretz, F., Hothorn, T. e Westfall, P. (2010). Confronti multipli con R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, BL (1994). Studio etologico dello sbadiglio nei primati. I. Analisi quantitativa e studio della causalità in due specie di scimmie del Vecchio Mondo (Cercocebus albigena e Macaca fascicularis). Etologia 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
Testo completo di CrossRef | Google Scholar
Dobson, A. J. (2002). Unintroduzione ai modelli lineari generalizzati, 2a edizione. Boca Raton, FL: Chapman and Hall.
Google Scholar
Katsuki, F. e Constantinidis, C . (2014). Attenzione bottom-up e top-down: processi differenti e sistemi neurali sovrapposti. Neuroscienziato 20, 509–521. doi: 10.1177 / 1073858413514136
Estratto di PubMed | Testo completo CrossRef | Google Scholar
Singer, T. (2006). Le basi neuronali e lontogenesi dellempatia e della lettura della mente: revisione della letteratura e implicazioni per la ricerca futura. Neurosci. Biobehav. Rev. 30, 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
Estratto di PubMed | Testo completo CrossRef | Google Scholar
Statistics Canada (2009). Categorie di età, raggruppamenti del ciclo di vita. Ottawa: Statistics Canada.
Google Scholar