Prelucrarea informațiilor este esențială în toate domeniile științei. În biologia moleculară, dogma centrală, inventată pentru prima dată de Francis Crick (Crick, 1958, 1970), este o coloană vertebrală clasică a celulelor vii pentru a executa în mod fundamental procese de la divizarea celulară la moarte prin căile de informații ADN, ARN și proteine. Mai precis, dogma centrală descrie transferul informațiilor de secvență în timpul replicării ADN, transcripției în ARN și translației în lanțuri de aminoacizi care formează proteine. În același timp, afirmă, de asemenea, că informațiile nu pot curge de la proteină la proteină sau acid nucleic.
De la apariția abordărilor sistemice și de mare randament în ultimele două decenii, acești pași largi, care nu includ detalii complexe de reglementare, au fost supuse unui control intens. Caracteristicile de reglementare lipsă, cum ar fi mecanismele de corectare / reparare a ADN-ului și îmbinarea alternativă a pre-ARNm, introduc mai mulți pași intermediari. Acești pași suplimentari interferează cu pașii cheie ai dogmei și probabil modifică dinamica informațiilor. În plus, epigenetica sau rolul jucat de structurile cromatinei, metilarea ADN-ului și modificările histonelor, par, de asemenea, să meargă împotriva căilor simple ale dogmei (Shapiro, 2009; Luco și colab., 2011). Splicing-ul proteinelor sau capacitatea unei proteine (inteine) de a-și modifica propria secvență, descoperite în vremurile recente (Volkmann și Mootz, 2012) și prioni, care modifică alte secvențe de proteine (Prusiner, 1998), ocolesc calea de transfer de informații a dogmă. Alte investigații au raportat erori sau nepotriviri între secvențele de ARN și ADN-ul lor de codificare (Hayden, 2011; Li și colab., 2011). Luate împreună, aceste date ridică îndoieli cu privire la validitatea dogmei centrale în contextul științei actuale și, prin urmare, pun sub semnul întrebării simplitatea fluxului liniar de informații (ADN către ARN și ARN către proteine).
Pentru a pune lucrurile în perspectivă, avem nevoie de instrumente analitice care să investigheze preocupările sau discrepanțele cu privire la teoria de lungă durată. O tehnică simplă, dar extrem de utilă pentru căutarea proprietăților globale în seturi de date cu randament ridicat este analiza statistică a corelației, care a fost utilizată pe scară largă și cu succes pentru a observa modele în sisteme complexe precum vremea (Stewart, 1990), piețele bursiere (Lo și MacKinlay , 1988) și cosmologie (Amati și colab., 2008). Există mai multe tipuri de analize de corelație care evaluează atât dependențele liniare (de exemplu, momentul produsului Pearson), cât și neliniare (de exemplu, rangul lui Spearman, informații reciproce) (Steuer și colab., 2002; Rosner, 2011). , analiza corelației produs-moment Pearson a devenit cea mai populară datorită capacității sale de a arăta structura organizațională în cea mai simplă formă.
În biologie, au existat numeroase lucrări care au studiat corelațiile în ARNm și date de expresie a proteinelor (vezi mai jos și tabelul 1). În teorie, atunci când sunt comparate două probe care conțin date de înaltă dimensiune (cum ar fi microarray și proteomic), analizele de corelație oferă o măsură a abaterii de la unitate ca sursă de diferență între probe Pe scurt, două eșantioane cu informații identice și complet neidentice vor afișa, respectiv, corelația unității (R2 = 1) și nulă (R2 = 0).
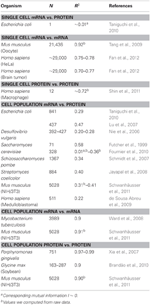
Tabelul 1. mRNA și proteine expr corelații de esență în diferite organisme.
Corelația perfectă (R2 = 1) este o situație idealizată care este departe de realitate, ca tehnică sau zgomotul experimental singur interferează și reduce corelația. Mai mult, ultimii ani au evidențiat existența zgomotului biologic: studiile asupra celulelor și moleculelor individuale au arătat stocasticitate în dinamica expresiei genice datorită efectului combinatorial al numerelor mici de copii moleculare și naturii cuantice a dinamicii promotorului (Raj și van Oudenaarden, 2009; Eldar și Elowitz, 2010). Pe de altă parte, populațiile clonale de celule prezintă eterogenitate la nivelurile unei expresii date de proteină per celulă în orice moment măsurat (Chang et al., 2008). Împreună, stochasticitatea și eterogenitatea sunt esențiale pentru producerea diversificării destinului celular, a variațiilor fenotipice și a amplificării semnalelor intracelulare (Locke și colab., 2011; Selvarajoo, 2012).
Fluctuațiile stochastice, sau zgomotul intrinsec, cauzează expresia unei specii moleculare să varieze în timp și între celule, ducând la răspunsuri necorelate (Elowitz și colab., 2002). Acest lucru este deosebit de important pentru ARNm și proteine cu un număr redus de copii. Astfel, corelația dintre eșantioane (celule) poate fi redusă datorită zgomotului intrinsec (Figura 1A). Alte surse de zgomot biologic datorate factorilor extrinseci includ variabilitatea dimensiunii celulei, numărul copiilor moleculare și fluctuațiile de mediu între celulele individuale.Acești factori distorsionează dogma centrală deterministă și probabil modifică corelațiile puternice în cele mai slabe (Figura 1B).
Un studiu recent a comparat expresiile ARNm și proteine ale Escherichia coli între celule individuale la nivel de moleculă unică și a oferit un scenariu care pune la îndoială dogma centrală. Taniguchi și colab. (2010) au dezvăluit că nu există nicio corelație (R2 ~ 0) între ARNm tufA individual și nivelurile de proteine în celule unice. În special, au ajuns la concluzia că lipsa corelației se datorează probabil diferențelor în durata de viață a ARNm și proteine. Deși aceasta este o explicație plauzibilă, Taniguchi și colab. au fost atenți să nu infirme ipoteza de lungă durată susținând că mediile în timp ale nivelurilor de ARNm ar trebui să se coreleze cu nivelurile de proteine. Cu toate acestea, nu s-au demonstrat dovezi care să demonstreze că acesta este cazul real și când am evaluat dependențele neliniare folosind informații reciproce (Steuer și colab., 2002; Tsuchiya și colab., 2010) în Taniguchi și colab. set de date, am constatat că rezultatul nu este dependent, adică I ~ 0. Acest lucru confirmă faptul că expresiile ARNm la proteine dintre celulele individuale la nivel de moleculă unică nu sunt în mod clar legate. Mai mult, atunci când se mărește la nivel de moleculă unică în graficul de corelație, este evident că corelațiile lor perechi sunt slabe (Figura 1A, inserare, pentru ilustrare).
În special, la nivelul populației celulare, Taniguchi et. al. au fost capabili să arate o corelație relativ mare între expresiile ARNm și proteine cu R2 = 0,29 (Figura 2A). De fapt, un alt studiu independent realizat de Lu și colab. (2007), pentru populația cu E. coli, au prezentat, de asemenea, o corelație relativ ridicată (R2 = 0,47). Analize similare efectuate pe Saccharomyces cerevisiae (Futcher și colab., 1999), fibrilastul NIH / 3T3 murin (Schwanhäusser și colab., 2011) și alte câteva populații celulare (Nie și colab., 2006; Schmidt și colab., 2007; Jayapal și colab. al., 2008; de Sousa Abreu și colab., 2009) toate au arătat structuri corelate între expresiile la nivel transcriptom și la nivel proteom (Tabelul 1). Deci, de ce nu există nicio corelație între expresiile ARNm individuale și proteinele din celule unice, în timp ce la nivel de populație se observă relații colective între expresiile ARNm la scară largă și expresiile proteinelor?
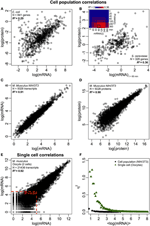
Figura 2. Corelații de expresie la nivel de Omics. Populații de celule: corelații ARNm-proteine în (A) E. coli (Taniguchi și colab., 2010) și (B) S. cerevisiae (Fournier și colab., 2010) între expresiile ARNm la t = 60 min și expresiile proteine la t = 360 min. Inserare: matricea de corelație între toate punctele de timp arată o creștere întârziată a corelațiilor dintre ARNm și proteine. (C) expresii mARN și (D) proteine între două probe de celule murine NIH / 3T3 (Schwanhäusser și colab., 2011). Celule unice: expresii ARNm (E) între două ovocite (Tang și colab., 2009). Liniile punctate roșii indică regiunile expresiilor ARNm scăzut (log (ARNm) < 5). (F) Zgomot (η2) versus log (expresii ARNm) pentru populația de celule (NIH / 3T3, puncte negre, Schwanhäusser și colab., 2011) și celule unice (ovocite, triunghiuri verzi, Tang și colab., 2009). Fiecare punct reprezintă valoarea pentru un grup de P = 100 mARN. η2 este aproape de zero pentru populația de celule pentru toate expresiile ARNm. Pentru celulele unice, η2 este cel mai mare pentru ARNm cu cele mai mici numere de copiere și se apropie de zero pentru numerele de copiere mai mari.
Credem există două motive majore pentru diferențe. În primul rând, așa cum sa menționat anterior, zgomotul, indiferent dacă este biologic sau non-biologic, reduce corelația. Deoarece analizele pe celule unice au arătat importanța stochasticității și variabilității, aceste efecte sunt cruciale pentru reducerea corelațiilor cu celule unice. La nivel de ansamblu, când celulele sunt eșantionate într-o populație, zgomotul total (intrinsec + extrinsec) este redus, deoarece zgomotul aleatoriu se anulează în toată gama de expresii moleculare (Figurile 1C-F), pentru a dezvălui răspunsul mediu și autoorganizarea ( Karsenti, 2008; Selvarajoo, 2011; Hekstra și Leibler, 2012; Selvarajoo și Giuliani, 2012). Prin urmare, apare un grad bun de corelație a expresiei mARN-proteină. În al doilea rând, pentru studiul cu o singură celulă (Taniguchi și colab., 2010), corelația individuală a expresiei mARN-proteină a fost comparată între numeroase celule. Cu toate acestea, în studiile privind populația celulară, comparația se face în întregime, între mii de mARN și proteine de mai multe ordine de mărime mai mari decât intervalul de expresie găsit pentru o singură moleculă între celule. Prin urmare, acest lucru duce la corelații mai mari la nivel de populație, deoarece efectul variațiilor moleculare unice devine neglijabil.
În ciuda structurilor corelate observate pentru populațiile de celule, există motive tangibile pentru abaterea mare de la corelația perfectă.După cum sa menționat anterior, un punct cheie este că mARN-urile și proteinele sunt localizate secvențial cu mai multe procese lipsă, nereprezentate în dogma centrală. Adăugarea intermediarilor lipsă de-a lungul unei căi biochimice va atrage o întârziere vizibilă a fluxului de informații (Selvarajoo, 2006, 2011; Piras și colab., 2011), iar corelația dintre aceștia ar putea suferi ca urmare. Acest lucru ar putea fi, de asemenea, o parte a faptului remarcat de Taniguchi și colab. că expresiile ARNm și proteine au durate de viață diferite. În special, această postulare este susținută într-o lucrare recentă pe S. cerevisiae tratată cu Rapamicină care a arătat că corelațiile temporale ale expresiei ARNm-proteine au fost inițial scăzute, R2 = 0,01 la 40 de minute, cu toate acestea, peste 360 de minute după perturbare, corelația a crescut, R2 = 0,36 (Fournier și colab., 2010, Figura 2B). Datele indică faptul că, la perturbarea chimică, răspunsul inițial dintre expresiile ARNm și proteine deviază din cauza întârzierii în timp și a diferitelor mecanisme cinetice dintre ele, precum și a efectelor secundare, cum ar fi interferența de semnalizare autocrină sau paracrină (Shvartsman și colab., 2002; Isalan și colab., 2008). Când efectele perturbației sunt atenuate în timp, a avut loc recuperarea corelațiilor.
Pentru a verifica în continuare postularea că procesele de întârziere secvențială sau durate diferite de viață sunt cruciale pentru scăderea corelațiilor mARN-proteină, am comparat R2 între aceeași specie moleculară a dogmei centrale (de exemplu, între ARNm și ARNm) în populațiile de celule și celule unice. Corelația expresiei mARN-transcriptom la nivel de expresie a ARNm între replicatele NIH / 3T3 (Schwanhäusser și colab., 2011) (Figura 2C) și Mycobacterium tuberculosis (Ward și colab., 2008) probele de populație celulară sunt ambele foarte mari, cu R2 > 0.9 (Tabelul 1). Astfel de corelații puternice sunt, de asemenea, observate între eșantioanele populației pentru expresiile proteine-proteine din celulele NIH / 3T3 (Schwanhäusser și colab., 2011) (Figura 2D), Porphyromonas gingivalis (Xia și colab., 2007) și Glicină max (Brandão și colab. , 2010) (Tabelul 1). Deoarece aceste date care compară aceleași specii produc corelații foarte mari, este de conceput că procesele de întârziere secvențială sau durate de viață diferite sunt responsabile pentru scăderea structurilor de corelație a nivelului populației dintre expresiile ARNm și proteine. Tang și colab., 2009), atunci când se compară expresii între ARNm-ARNm, se observă o structură foarte corelată (R2 = 0,92, Figura 2E). Cu toate acestea, concentrându-ne doar pe ARNm cu expresie redusă (cu expresii logaritmice < 5), zgomotul stochastic reduce corelația pe perechi destul de dramatic (R2 < 0,54). Pentru a testa acest rezultat, am evaluat zgomotul, η2 = σ2XY / μ2XY, pe întreaga expresie a ARNm (Figura 2F). Am observat că η2 este cel mai mare pentru cele mai mici expresii, datorită efectului pronunțat al fluctuațiilor stochastice în comparație cu expresiile lor și se apropie de zero pentru expresiile superioare, unde un astfel de zgomot devine mai puțin semnificativ (Piras și colab., 2012). Pentru populația celulară, așa cum era de așteptat, se observă zgomot aproape zero pe întregul interval de expresie datorită eliminării zgomotului aleatoriu (Figurile 1E, F).
Au fost, de asemenea, structuri foarte corelate pentru întreaga expresie mARN-mARN. raportate pentru celule canceroase unice (Fan și colab., 2012), deși mai puțin semnificative cu R2 ~ 0,7 (Tabelul 1). Mai mult, compararea expresiilor proteină-proteină în macrofagele umane stimulate de LPS au arătat, de asemenea, corelații mari, R2 ~ 0,72 (Shin și colab., 2011) (Tabelul 1). Deși nu există nicio corelație între expresiile individuale ale ARNm-proteine în celule unice, corelația la scară largă sau la nivel omic între aceleași specii moleculare din celule individuale este foarte mare. , datele la nivelul omicilor indică faptul că corelațiile dintre aceleași specii moleculare (ARNm față de ARNm și proteine față de proteine) sunt semnificativ mai mari decât între diferite specii (ARNm față de proteine). Acest lucru reflectă faptul că, deși procesele de întârziere și durate diferite de viață sunt cheia pentru reducerea corelațiilor, aceste mecanisme nu sunt suficiente pentru a susține lipsa structurii de corelație observată între transcrierea individuală a celulelor individuale și expresiile proteinelor.
până acum, prin investigarea expresiilor la scară largă ale ARNm și proteinelor din diferite sisteme celulare, am arătat că structurile de corelație apar la scară globală. Cu toate acestea, analizele de corelație relevă doar conectivitatea dintre două probe testate și nu arată direcția fluxul de informații. Pentru ca dogma centrală să fie valabilă la scară globală, fluxul general de informații ar trebui să fie de la ADN la proteine. Un astfel de flux de informații a fost demonstrat de nenumărate alte studii care implică perturbarea receptorilor populațiilor de celule și monitorizarea rezultatului dinamica factorilor de transcripție care se leagă de ADN și inducerea expresiilor genei la scară largă (Figura 3A).De exemplu, în cazul celulelor imune stimulate de LPS, s-a demonstrat că activarea factorului de transcripție NF-κB are loc în jur de 15 minute (Liu și colab., 1999), inducerea genelor sale în aval la aproximativ 30 min (Liu și colab., 1999; Xaus și colab., 2000; Selvarajoo și colab., 2008) și traducerea proteinelor corespunzătoare în regiunea de 60-90 min (Kawai și colab., 1999; Xaus și colab. ., 2000) (Figura 3B). O astfel de direcție secvențială a transcrierii globale la fluxul de informații de traducere este, de asemenea, observată pentru sistemele bacteriene, cum ar fi E. coli, la nivel de populație celulară (Golding și colab., 2005).

Figura 3. Fluxul de informații al dogmei centrale. (A) Schema expresiei TNF indusă de LPS / TLR4, prin factorul de transcripție NF-κB și gena tnf, după fluxul de informații liniar. (B) Profiluri experimentale temporale ale activității de legare a promotorului expresiilor NF-κB (panouri superioare), tnf (panouri medii) și TNF (panouri inferioare) la nivelul populației celulare. (C) Profiluri temporale schematice ale dinamicii promotorului, ARNm și expresii proteice la nivel unicelular (Raj și van Oudenaarden, 2009).
În mod alternativ, investigațiile la rezoluția unei singure celule relevă fluctuații aleatorii asupra fluxului liniar de informații: factorii de transcripție care se leagă de regiunile promotorului ADN sunt cuantice, rezultând comportamentul de explozie al transcripției ARNm și, ulterior, induce variabilitatea în traducerea proteinelor, chiar și între celule identice (Figura 3C) (Raj și van Oudenaarden, 2009; Eldar și Elowitz, 2010; Locke și colab., 2011; Hekstra și Leibler, 2012; Selvarajoo, 2012). Ca rezultat, în orice moment particular, răspunsul molecular individual pentru celule individuale este destul de zgomotos comparativ cu scara medie a populației (Selvarajoo, 2011).
Concluzii
Exemplele prezentate în această lucrare evidențiază diferențele în ordinea valorilor de corelație observate între speciile din dogma centrală asupra populațiilor celulare și celulelor unice. Analizele statistice din populațiile de celule ilustrează că corelația expresiei dintre aceeași specie moleculară este foarte mare și între specii este moderat de mare. Deși corelațiile cu o singură celulă între aceleași specii sunt comparabile cu populațiile de celule, acestea au arătat o dispersie mai largă în graficele lor de expresie datorită efectului pronunțat al zgomotului biologic, în special pentru transcrierile cu un număr mic de copii. În special, corelația în perechi a celulelor unice devine zero pentru moleculele individuale (Taniguchi și colab., 2010). De fapt, fluctuațiile stochastice și variabilitatea expresiilor moleculare sunt cunoscute ca fiind funcționale în generarea deciziei destinului celulei și în înclinarea stărilor celulare (Losick și Desplan, 2008; Eldar și Elowitz, 2010; Kuwahara și Schwartz, 2012). Credem că corelațiile puternice la nivel omic apar ca urmare a rețelelor de reglare strânse a genelor și proteinelor în mii de molecule (Barabási și Oltvai, 2004; Karsenti , 2008) rezultând răspunsuri medii emergente. Analizând un număr mic sau molecule individuale, structura de corelație nu poate fi observată.
În general, este de conceput că vizualizarea fluxului de informații al ADN-ului unic către proteină va pune sub semnul întrebării dogma centrală deoarece răspunsul fiecărei molecule la un moment dat probabil nu se va corela. Cu toate acestea, la nivel global, observarea răspunsului determinist mediu sugerează că echilibrul net al genetei informațiile ic rămân în extrema dreaptă a căilor. Prin urmare, dogma centrală ar trebui privită ca un flux de informații celular macroscopic pe o scară omică la scară largă, și nu la o singură genă la nivel de proteină. Ca atare, credem că simplitatea sa va rămâne în continuare ca unul dintre cei mai influenți stâlpi teoretici ai sistemelor vii.
Declarația privind conflictul de interese
Autorii declară că cercetarea a fost realizată în absența oricărei relații comerciale sau financiare care ar putea fi interpretată ca un potențial conflict de interese.
Mulțumiri
Kentaro Hayashi este mulțumit pentru comentarii. Fondul de cercetare al orașului Tsuruoka și al prefecturii Yamagata este apreciat pentru sprijinul lor.
Crick, F. (1958). Despre sinteza proteinelor. Simp. Soc. Exp. Biol. 12, 139-163.
Rezumat Pubmed | Text complet publicat
Crick, F. (1970). Dogma centrală a biologiei moleculare. Nature 227, 561-563.
Pubmed Abstract | Text complet publicat
Hayden, E. C. (2011). Dovezi ale modificării ARN stârnește dezbaterea. Nature 473, 432.
Pubmed Abstract | Text complet publicat | CrossRef Full Text
Hekstra, D. R. și Leibler, S. (2012). Contingența și legile statistice în replicarea ecosistemelor microbiene închise. Cell 149, 1164–1173.
Pubmed Abstract | Text complet publicat | CrossRef Full Text
Kuwahara, H. și Schwartz, R. (2012).Câștig în stare de echilibru stochastic într-un proces de expresie genică cu control al degradării ARNm. J. R. Soc. Interfață 9, 1589-1598.
Pubmed Abstract | Text complet publicat | CrossRef Text complet
Nie, L., Wu, G. și Zhang, W. (2006). Corelația expresiei ARNm și a abundenței de proteine afectate de caracteristicile secvenței multiple legate de eficiența translațională în Desulfovibrio vulgaris: o analiză cantitativă. Genetica 174, 2229-2243.
Pubmed Abstract | Text complet publicat | CrossRef Text complet
Prusiner, S. B. (1998). Prioni. Proc. Natl. Acad. Știință. S.U.A. 95, 13363–13383.
Pubmed Abstract | Text complet publicat | CrossRef Text complet
Rosner, B. (2011). Bazele biostatisticii. Al 7-lea Edn. Boston, MA: Duxbury Press.
Selvarajoo, K. (2006). Descoperirea mașinilor de activare diferențială a căilor de semnalizare a receptorului cu taxă 4 în eliminările MyD88. FEBS Lett. 580, 1457–1464.
Pubmed Abstract | Text complet publicat | CrossRef Text complet
Selvarajoo, K. (2011). Legea macroscopică a conservării a dezvăluit în dinamica populației de semnalizare a receptorilor asemănători. Cell Commun. Semnal. 9, 9.
Rezumat Pubmed | Text complet publicat | CrossRef Text complet
Selvarajoo, K. (2012). Înțelegerea deciziilor biologice multimodale din dinamica unei singure celule și a populației. Wiley Interdiscip. Rev. Syst. Biol. Med. 4, 385-399.
Rezumat Pubmed | Text complet publicat | CrossRef Text complet
Stewart, T. R. (1990). O descompunere a coeficientului de corelație și utilizarea acestuia în analiza abilităților de prognozare. Prognoza meteo. 5, 661-666.