Introduction
Le bâillement est une séquence involontaire douverture de la bouche, dinspiration profonde, dapnée brève et dexpiration plus ou moins lente (Baenninger, 1997; Walusinski et Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Lorsquil est suscité, un bâillement ne peut pas être totalement supprimé. Par conséquent, il a été défini comme un modèle stéréotypé ou de type réflexe (Lehmann, 1979; Provine, 1986). Chez Homo sapiens, plusieurs hypothèses ont été avancées avec un support variable pour expliquer les mécanismes et les fonctions du bâillement spontané, comme loxygénation (fonction respiratoire causée par lhypoxie), le comportement lié au stress (causé par lexcitation) ou la thermorégulation (causée par lhyperthermie); Guggisberg et al., 2010; Massen et al., 2014; Gallup et Gallup, 2019). Étant une réponse physiologique, le bâillement peut être affecté par des facteurs internes et externes tels que lheure de la journée (Giganti et Zilli, 2011) ou la température intracrânienne / cérébrale (Gallup et Eldakar, 2013).
Le bâillement peut être autonome et / ou présenté aux autres (Moyaho et al., 2017; Palagi et al., 2019) Chez les primates humains et non humains, selon les espèces, lorsque le bâillement est montré aux autres, il peut communiquer une menace (Troisi et al., 1990; Delegte, 1994) et / ou des changements physiologiques et comportementaux (Provine et al., 1987 ; Leone et al., 2015; Zannella et al., 2015). Chez lhomme, le bâillement est une réponse modulée socialement car il peut être inhibé par une présence sociale réelle – et non virtuelle – (Gallup et al., 2019) et parce quun bâillement peut être déclenché par le bâillement de quelquun dautre, à la suite dun phénomène connu comme bâillements contagieux (Provine, 1989, 2005). La contagion du bâillement peut être provoquée même si le bâillement est entendu mais pas vu (Arnott et al., 2009; Massen et al., 2015).
Chez les humains, leur espèce de singe phylogénétiquement la plus proche (chimpanzés: Pan paniscus ; bonobos: Pan troglodytes) et le singe africain Theropithecus gelada, le bâillement contagieux nest pas seulement présent (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; mais voir: Amici et al., 2014) mais aussi modulée socialement parce que la réponse bâillante est la plus élevée lorsque certaines catégories dindividus sont impliquées (par exemple, parents, membres du groupe, dominants; Palagi et al., 2009; Campbell et de Waal, 2011, 2014; Norscia et Palagi, 2011; Demuru et Palagi, 2012; Massen et al., 2012). Deux arguments principaux ont été présentés pour expliquer cette asymétrie sociale dans le bâillement contagieux, qui ont été regroupés en deux hypothèses principales: lhypothèse du biais émotionnel (EBH), reliant le bâillement contagieux au transfert émotionnel, et lhypothèse du biais attentionnel (ABH), qui considère le bâillement contagieux en tant que réponse motrice sujette à des différences dans les processus attentionnels descendants (Palagi et al., 2020).
LEBH prédit que lasymétrie sociale observée dans les taux de contagion du bâillement reflète des différences dans les le lien social, un proxy du lien affectif, entre les individus. Cette hypothèse est étayée par des preuves que les taux de contagion du bâillement suivent une tendance empathique (sensu Preston et de Waal, 2002), étant les plus élevés entre les individus partageant un lien émotionnel le plus fort. Plus précisément, Norscia et Palagi (2011) ont constaté que chez les humains, les taux de contagion bâillonnés sont plus élevés en réponse aux parents et amis quen réponse aux connaissances et aux étrangers. Chez les chimpanzés adultes, la contagion du bâillement est plus élevée entre les membres du groupe par rapport aux membres de lextérieur du groupe (Campbell et de Waal, 2011) et chez les bonobos, les taux de contagion du bâillement sont les plus élevés entre les individus qui saffilient davantage entre eux (Demuru et Palagi, 2012). Dans une enquête comparative incluant à la fois des humains et des bonobos, Palagi et al. (2014) ont constaté que les taux de contagion du bâillement étaient plus affectés par la qualité des relations entre les individus que par les espèces auxquelles les sujets appartenaient. De plus, chez les humains, la contagion du bâillement augmente avec lâge lorsque la capacité didentifier les émotions des autres augmente et diminue avec la vieillesse lorsque cette capacité diminue (Wiggers et van Lieshout, 1985; Anderson et Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen et Anderson, 2011; Bartholomew et Cirulli, 2014). Les taux de contagion du bâillement augmentent de la petite enfance à lâge adulte également chez les chimpanzés (Madsen et Persson, 2013).
LABH prédit que lasymétrie sociale observée dans la contagion du bâillement peut être due à des différences dattention sociale et visuelle (Massen et Gallup, 2017). En particulier, les niveaux les plus élevés de bâillement contagieux seraient dus à lattention visuelle sélective et descendante supplémentaire accordée aux individus qui sont plus pertinents pour lobservateur, tels que les sujets familiers, comme cela se produit chez les humains et les geladas, ou les dominants, car il se produit chez les chimpanzés ou les bonobos (Yoon et Tennie, 2010; Massen et al., 2012; Massen et Gallup, 2017).Selon Massen et Gallup (2017), lABH serait étayée par les preuves existantes sur la détection visuelle différente et lencodage perceptif visuel des visages de sujets familiers et / ou en groupe par rapport à ceux inconnus (par exemple, Buttle et Raymond, 2003 ; Ganel et Goshen-Gottstein, 2004; Jackson et Raymond, 2006; Michel et al., 2006).
Dans cette étude, nous avons analysé des données sur le bâillement recueillies pendant 9 ans sur des humains dans leur milieu naturel et nous avons extrapolé les cas dans lesquels le bâillement émis par un sujet pouvait être entendu mais non vu par un répondant potentiel (bâillement auditif). En considérant uniquement les cas dans lesquels le signal visuel du stimulus bâillement nétait pas détectable, nous avons vérifié si lasymétrie sociale précédemment observée dans les taux de contagion bâillante persistait ou non. En particulier, nous avons testé les prédictions alternatives suivantes dérivées des deux hypothèses présentées ci-dessus (EBH et ABH).
Prédiction 1a: selon lEBH, les taux de contagion du bâillement sont influencés par la force de linter -un lien social individuel – un proxy du lien émotionnel – en soi et non par une attention visuelle sélective descendante différente accordée à certains individus en particulier. Si cette hypothèse est étayée, nous nous attendons à observer le biais social également lorsque le signal visuel du stimulus bâillement est exclu et que les taux de bâillement auditif contagieux seront plus élevés entre les individus fortement liés par rapport aux individus faiblement liés.
Prédiction 1b: selon lABH, les niveaux plus élevés de contagion de bâillement entre les individus fortement liés par rapport aux individus faiblement liés seraient liés à lattention visuelle sélective descendante la plus proche que les individus accordent aux individus qui sont pertinents pour eux, par exemple, la famille et copains. Si cette hypothèse est confirmée, le biais social observé dans la contagion du bâillement devrait disparaître lorsque seuls les bâillements auditifs sont pris en compte car le signal visuel ne peut être assisté par le répondant potentiel.
Matériel et méthodes
Collecte de données et définitions opérationnelles
Pour cette étude, nous avons considéré les bâillements vocalisés émis par un sujet qui ne pouvait être entendu – mais pas vu – par un répondant potentiel (ci-après: bâillements auditifs). Lémetteur et les répondeurs potentiels devaient être dans une plage ≤ 5 m. Les bâillements vocaux impliquaient lutilisation de cordes vocales et les bâillements qui nimpliquaient quune forte inspiration / expiration nétaient pas considérés comme vocalisés; 294 cas de bâillements auditifs ont été extrapolés à partir dun ensemble de données dun total de 2001 épisodes de bâillements recueillis sur 9 ans – de 2010 à 2019 – en utilisant la méthode déchantillonnage de toutes les occurrences (Altmann, 1974). Plus précisément, des bâillements auditifs ont été recueillis de novembre 2010 à mai 2019, de 05h30 à 02h30, sur des sujets humains caucasiens, âgés de 18 à 77 ans, au cours de leurs activités habituelles, par exemple sur les lieux de travail, au cours des repas, lors de réunions sociales, etc. ., les sujets ignorant quils ont été observés et en labsence de toute source externe évidente danxiété. La base de données de bâillement auditif comprenait 193 dyades répondeurs potentiels de lacet. Selon la situation, les informations ont été enregistrées, non notées, au moyen de codes alphanumériques et saisies directement dans des feuilles de calcul, tapées sur téléphone portable ou écrites sur papier, puis entrées dans des feuilles de calcul pour élaboration ultérieure. Des informations de base telles que lâge et la relation entre les personnes étaient connues des auteurs. Les répondeurs potentiels étaient codés comme étant en état de non-vue lorsque leur tête était tournée de 180 ° par rapport à la gâchette ou lorsquun obstacle physique bloquant la vue était présent empêchant le répondant potentiel de voir le visage et le corps de la gâchette. Le déclencheur et le répondeur nont jamais été complètement isolés (par exemple, dans deux pièces séparées avec des portes fermées) lun de lautre. La proximité sociale a été recueillie à quatre niveaux: 0 = étrangers, qui ne sétaient jamais rencontrés auparavant; 1 = connaissances, qui partageaient exclusivement une relation indirecte basée sur un troisième élément extérieur, à savoir le devoir de travail (collègues) ou des amis en commun (amis damis); 2 = amis, individus non apparentés partageant une relation directe non exclusivement liée à un troisième élément externe; 3 = partenaires réguliers et parents (r ≥ 0,25). La littérature précédente rapporte que les réponses de bâillement peuvent être déclenchées dans les 5 minutes après avoir regardé le bâillement de quelquun dautre (le bâillement de la gâchette) (Provine, 1986), avec un maximum dans la première minute (Provine, 2005; Palagi et al., 2014). La littérature rapporte également quà partir de la quatrième minute, la probabilité dautocorrélation est la plus élevée (ce qui signifie que la présence dun bâillement effectué par un sujet à t0 augmente la probabilité davoir un autre bâillement par le même sujet à t (0 + X) où X est lunité de temps croissante; Kapitány et Nielsen, 2017). Par conséquent, nous avons considéré les réponses de bâillement se produisant dans une fenêtre de temps de 3 minutes à partir du bâillement émis par la gâchette.Pour réduire davantage le biais dautocorrélation, en cas de chaîne de bâillements émis par la gâchette (plus de bâillements émis dans la fenêtre de temps de 3 min), nous avons enregistré comme réponse seulement le premier bâillement effectué après la perception du dernier bâillement. Nous avons codé un bâillement comme « spontané » alors quaucun autre sujet navait bâillé dans les 5 minutes précédant lévénement de bâillement.
Analyses statistiques
Pour les analyses, les variables suivantes ont été prises en compte: occurrence de contagion, codé comme suit: 1 = présence, 0 = absence; le lien social a été entré avec les quatre niveaux définis ci-dessus (0 = étrangers; 1 = connaissances; 2 = amis; 3 = parenté); le sexe du déclencheur et de lobservateur a été étiqueté comme : M = homme, F = femme; les classes dâge du déclencheur et du répondant ont été codées comme suit: yo = jeune (18–24 ans); ad = adulte (25–64 ans); se = senior (ci-dessus 65 ans) (Statistique Canada, 2009); les plages horaires étaient codées comme suit: 1 = 5 h 30 à 09 h 00; 2 = 9 h 01 à 12 h 30; 3 = 12 h 31 à 16: 00 h 00; 16 h 00 à 19 h 30; 5 = 19 h 31 à 23 h 00; 6 = 23 h 01 à 02 h 30 (Giganti et Zilli, 2011). La base de données (voir la fiche technique supplémentaire) comprenait 84 hommes, 69 femmes, 16 jeunes (yo), 122 adultes (ad) et 15 seniors (SE). Pour tester si loccurrence du bâillement la contagion était influencée par le lien des facteurs (0 = étrangers; 1 = connaissances; 2 = amis; 3 = kin), le sexe du déclencheur (Trigger_sex), le sexe du répondant (Responder_sex), la classe dâge du déclencheur (Trigger_ageclass) et la classe dâge du répondant (Responder_ageclass) et la tranche horaire (de 1 à 6 ), nous avons utilisé un modèle mixte linéaire généralisé (GLMM) qui incluait ces cinq prédicteurs en tant queffets fixes et les identités de déclencheurs (déclencheurs) et de répondeurs (répondeurs) comme effets aléatoires. Nous avons installé les modèles dans R (R Core Team, 2018; version 3.5.1) en utilisant la fonction lmer du R-package lme4 (Bates et al., 2015). Nous avons établi la signification du modèle complet par rapport à un modèle nul ne comprenant que les effets aléatoires (Forstmeier et Schielzeth, 2011). Nous avons utilisé un test du rapport de vraisemblance (Dobson, 2002) pour tester cette signification (ANOVA avec largument «Chisq»). Nous avons calculé les valeurs de p pour les prédicteurs individuels sur la base des tests du rapport de vraisemblance entre le modèle complet et le modèle nul en utilisant le -fonction « drop1 » (Barr et al., 2013). La variable de réponse étant binaire, nous avons utilisé une distribution derreur binomiale. Nous avons testé si linteraction entre les sexes ou les classes dâge du déclencheur et du répondant était significative, mais comme elles ne létaient pas, nous ne les avons pas incluses dans le modèle. Nous avons utilisé un package de contraste multiple (multcomp) pour effectuer toutes les comparaisons par paires pour chaque niveau de liaison avec le test de Tukey (Bretz et al., 2010). Nous avons signalé les valeurs p, lestimation (Est), lerreur standard (SE) et les valeurs z ajustées par Bonferroni.
Résultats

Figure 1. Graphique linéaire de leffet du lien social entre le déclencheur et le répondeur (axe X) sur loccurrence moyenne de contagion acoustique du bâillement (axe Y). Les amis et les parents affichent des fréquences de contagion de bâillement nettement plus élevées que les étrangers et les connaissances (test de Tukey: amis contre étrangers p < 0,001; parents contre étrangers p < 0,001; amis contre connaissances p < 0,001; parents contre connaissances p < 0,001; autres combinaisons, ns).
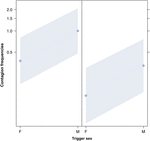
Figure 2. Graphique linéaire de leffet du sexe du déclencheur (axe X) sur loccurrence moyenne de contagion acoustique en bâillement (axe Y). Effet du sexe déclencheur sur la contagion acoustique du bâillement lorsque le répondeur est une femme (à droite) et un homme (à gauche). Les bâillements des mâles provoquent plus de bâillements que ceux des femelles quel que soit le sexe du répondant (résultat du GLMM, p = 0,022). Les points représentent leffet de la variable de réponse sur la variable indépendante «sexe du déclencheur», en fonction de la valeur prédite par le modèle. Les bandes colorées indiquent lintervalle de confiance à 95%.
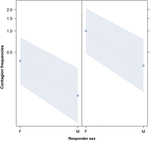
Figure 3. Diagramme linéaire de leffet du sexe du répondeur (axe X) sur loccurrence moyenne de contagion acoustique du bâillement (axe Y). Effet du sexe du répondeur sur la contagion acoustique du bâillement lorsque la gâchette est une femme (droite) ou un homme (gauche). Les femmes répondent beaucoup plus que les hommes. du sexe du déclencheur (résultat du GLMM, p = 0,021). Les points représentent leffet de la variable de réponse sur la variable indépendante «sexe du répondant», sur la base de la valeur prédite par le modèle. Les bandes colorées montrent lintervalle de confiance à 95%.
Discussion
Cette étude montre pour la première fois que la contagion du bâillement est significativement affectée par le lien social entre individus (Tableau 1) même lorsque les stimuli de déclenchement sont des bâillements auditifs, que nous avons définis comme des bâillements vocalisés qui pouvaient être entendus mais non vus (repère visuel indétectable, repère auditif détectable). En particulier, les bâillements auditifs contagieux sont significativement plus fréquents entre parents et amis quentre étrangers et connaissances (Figure 1). Cette découverte soutient la prédiction 1a basée sur lEBH et non la prédiction 1b basée sur lABH, conduisant à la conclusion que chez lhomme, lattention visuelle sélective de haut en bas ne peut pas être le principal moteur de lasymétrie sociale observée dans les taux de contagion du bâillement (Norscia et Palagi , 2011; Norscia et al., 2016a). De plus, le sexe du déclencheur et le sexe du receveur avaient un effet significatif sur les taux de contagion du bâillement, les hommes – en tant que déclencheurs – auxquels les autres répondaient plus fréquemment que les femmes (Figure 2) et les femmes répondant plus fréquemment aux bâillements des autres que les hommes (Figure 3).
Contrairement à Bartholomew et Cirulli (2014), nous navons trouvé aucun effet de lâge sur la contagion du bâillement, très probablement parce que notre base de données sur les bâillements auditifs avait une forte prévalence dadultes (25-64 ans vieille). Les niveaux les plus élevés de contagion auditive du bâillement chez les femmes par rapport aux hommes confirment le biais de genre observé dans des conditions naturalistes chez les humains susceptibles de contagion par le bâillement par Norscia et al. (2016a, b) lors de lexamen dun ensemble de données plus large qui comprenait également des bâillements qui pourraient être vus par le répondant potentiel (avec une modalité sensorielle bâillante – vision, audition ou les deux – naffectant pas la réponse). Le biais de genre est également en accord partiel avec les résultats précédents obtenus dans des contextes contrôlés, y compris le signal visuel (Chan et Tseng, 2017; mais voir Norscia et Palagi, 2011; Bartholomew et Cirulli, 2014). On a émis lhypothèse que le degré élevé de contagion du bâillement chez les femmes pourrait informer la contagion émotionnelle (Norscia et al., 2016a), à la lumière des capacités empathiques apparemment plus élevées – liées à la maternité – des femmes que des hommes (Christov-Moore et al. ., 2014). Cependant, cette question est toujours en débat car les différences culturelles entre les sociétés humaines peuvent modeler les dynamiques de liens sociaux dune manière différente. Il est donc compliqué, à ce stade de la connaissance, de démêler les facteurs culturels, la qualité des relations interpersonnelles et linfluence du genre dans la distribution de la contagion du bâillement. Nos résultats montrent également que les hommes fonctionnent mieux que les femmes comme déclencheurs et l’hypothèse la plus parcimonieuse à ce sujet pourrait être que les vocalisations des hommes peuvent être mieux entendues dans des milieux naturels, souvent caractérisés par des bruits de fond. En effet, la perception du genre de la voix repose principalement sur la fréquence fondamentale qui est en moyenne plus basse dune octave chez les voix masculines que féminines, les vocalisations à basse fréquence voyageant plus loin que celles à haute fréquence (Marten et Marler, 1977; Latinus et Taylor, 2012) . Cependant, à notre connaissance, il ny a pas détude spécifique traitant du possible biais de genre dans laudibilité du bâillement et une enquête plus approfondie avec des essais expérimentaux en condition contrôlée est donc nécessaire pour vérifier cette spéculation.
Dans cette étude, nous avons également trouvé que les différences dans les taux de contagion du bâillement entre les catégories (famille et amis, étrangers et connaissances) ne peuvent pas être expliquées par des différences dans lattention visuelle sélective descendante. Cette constatation est conforme à la littérature précédente. Le bâillement contagieux semble impliquer des zones du cerveau qui sont plus liées au réseau orienté de bas en haut que des zones liées de haut en bas. De plus, la contagion du bâillement nest ni sensible aux signaux sensoriels présents dans le signal (auditif, visuel ou audiovisuel) (Arnott et al., 2009; Norscia et Palagi, 2011) ni affectée par la perspective visuelle du stimulus déclencheur ( les bâillements dans des orientations de 90 °, 180 ° et 270 ° sont capables de déclencher des réponses de bâillement sous forme de bâillements frontaux à 0 °; Provine, 1989, 1996). Chan et Tseng (2017) ont constaté que la capacité de détecter un bâillement en tant que tel (sensibilité de détection perceptuelle) était liée à la durée du regard sur les yeux du visage libérant le stimulus, mais les modèles de regard ne pouvaient pas moduler le bâillement contagieux. Chez les chimpanzés, les fréquences de bâillements contagieux étaient les plus élevées entre le même groupe que les individus de différents groupes, même si les répondants ont regardé plus longtemps les vidéos de chimpanzés hors groupe (Campbell et de Waal, 2011). Largument selon lequel lattention sélective visuelle peut biaiser les taux de contagion du bâillement dans une direction spécifique (les sujets répondant plus à la famille que les étrangers) est également miné par labsence de tout modèle spécifique dattention sociale chez les primates humains et non humains. Par eye-tracking (appliqué pour mesurer le temps de visionnage) et en montrant des visages inconnus à leurs sujets expérimentaux, Méary et al.(2014) ont observé que les humains étaient orientés vers les visages de leur propre race alors que lattention des macaques rhésus était plus attirée par les nouveaux visages que par les mêmes visages. Kawakami et coll. (2014) ont observé que les sujets humains accordaient plus dattention aux yeux des membres du groupe ethnique et au nez et à la bouche des membres du groupe ethnique. La même étude a également révélé que lattention visuelle ne dépendait pas de la race cible. En mesurant combien de temps les sujets expérimentaux ont regardé lécran, Whitehouse et al. (2016) ont observé que les macaques de Barbarie accordaient plus dattention aux vidéos de grattage de non-étrangers que dindividus étrangers, mais ont également noté que parmi les non-étrangers, les macaques accordaient le plus dattention aux personnes avec lesquelles ils partageaient un lien social faible. En mesurant les taux de regard, Schino et Sciarretta (2016) ont observé que les mandrills regardaient plus leurs propres parents que les non-parents, mais aussi plus les partenaires dominants que subordonnés du groupe. Par conséquent, ces études (utilisées pour soutenir lABH) ne décrivent aucun modèle unique dattention sélective. Un autre point important à considérer est la définition même de la familiarité et de lappartenance à un groupe adoptée par la plupart des études utilisées pour soutenir lABH (Massen et Gallup, 2017). Ces études ont montré une meilleure détection visuelle et un codage perceptif visuel des visages de sujets familiers / dans le groupe par rapport à ceux qui ne le sont pas, mais ont défini la familiarité et lappartenance au groupe non sur la base des relations personnelles entre les individus. Au lieu de cela, la familiarité ou lappartenance à un groupe a été définie sur la base de connaissances indirectes (par exemple, photo de personnes célèbres ou dun sujet déjà montré dans les phases préalables au procès) ou de race commune (par exemple, Buttle et Raymond, 2003; Ganel et Goshen- Gottstein, 2004; Jackson et Raymond, 2006; Michel et al., 2006). Cette définition convient aux fins de ces études mais elle lest moins si les résultats sont utilisés pour proposer des explications alternatives de linfluence que le lien social réel – basé sur des relations réelles – peut avoir sur un phénomène, en loccurrence la contagion bâillante . Par exemple, Michel et al. (2006) ont observé que les sujets caucasiens et asiatiques pouvaient mieux reconnaître les visages de même race, mais cette différence nétait pas présente chez les sujets asiatiques qui vivaient depuis environ un an chez les Caucasiens. Un autre point de discussion concerne la présence de contagion du bâillement chez les enfants atteints de troubles du spectre autistique (TSA), qui montrent fréquemment des altérations de lattention visuelle (Richard et Lajiness-O’Neill, 2015). Chez les enfants atteints de TSA, la contagion du bâillement peut être absente (Senju et al., 2007), altérée (Helt et al., 2010) ou similaire aux enfants en développement typique lorsque les sujets sont amenés à rediriger leur attention vers le stimulus vidéo pendant les essais expérimentaux. (Usui et al., 2013). Dans une étude récente, Mariscal et al. (2019) ont constaté que la contagion du bâillement chez les enfants atteints de TSA était positivement liée à la concentration sanguine docytocine, lhormone impliquée dans lattachement parental et social (Decety et al., 2016) et ont émis lhypothèse que la contagion du bâillement chez les enfants atteints de TSA pourrait être liée à une moyenne variable. concentrations docytocine dans différentes cohortes détudes (Mariscal et al., 2019). Cette constatation est conforme à lhypothèse EBH qui lie les taux de contagion du bâillement au lien social, qui peut refléter un lien émotionnel.
Conclusion
Notre étude ajoute à la discussion sur les mécanismes sous-jacents au asymétrie sociale dans la contagion du bâillement (pour une critique critique: voir Adriaense et al., 2020; Palagi et al., 2020), en montrant que la contagion du bâillement est probablement associée à une attention sélective ascendante plutôt que descendante. Lattention ascendante est principalement dirigée par la perception sensorielle du stimulus déclencheur, tandis que lattention sélective descendante est un processus volontaire et soutenu dans lequel un élément particulier est sélectionné en interne et focalisé ou examiné (Katsuki et Constantinidis, 2014). À cet égard, le stimulus acoustique (bâillement auditif) émis par la gâchette a été entendu et pouvait provoquer une réponse bâillante dans le récepteur, même si le récepteur ne prêtait aucune attention visuelle volontaire à la gâchette. De plus, les taux de réponse au bâillement étaient modulés socialement, la contagion auditive du bâillement étant la plus élevée chez les individus les plus fortement liés les uns aux autres. Par conséquent, lattention sélective descendante nest pas le principal moteur de lasymétrie sociale observée dans la contagion du bâillement, qui semble être un phénomène induit par un stimulus lié aux processus dattention ascendante. Une enquête plus approfondie est nécessaire pour comprendre si et de quelle manière dautres formes dattention ou étapes pré-attentives peuvent affecter la contagion du bâillement.
Déclaration de disponibilité des données
Lensemble de données utilisé pour cette étude est fourni dans le Matériel supplémentaire.
Déclaration déthique
Létude impliquant des participants humains a été revue et approuvée par le Comitato di Bioetica dAteneo (Comité de bioéthique de lUniversité) – Université de Turin (réf n ° 451945).Le consentement éclairé écrit pour la participation nétait pas requis pour cette étude conformément à la législation nationale et aux exigences institutionnelles.
Contributions des auteurs
IN et EP ont procédé à la collecte de données et conçu et rédigé le manuscrit. AZ a aidé à la collecte de données et à la révision des manuscrits, des figures et des tableaux. MG a effectué des analyses statistiques et rédigé la partie correspondante du manuscrit.
Conflit dintérêts
Les auteurs déclarent que la recherche a été menée en labsence de toute relation commerciale ou financière qui pourrait être interprété comme un conflit dintérêts potentiel.
Remerciements
Les auteurs tiennent à remercier le professeur Roberto Barbuti et le professeur Cristina Giacoma davoir soutenu la recherche à lUniversité de Pise et Université de Turin, respectivement.
Matériel supplémentaire
Altmann, J. (1974). Etude observationnelle du comportement: méthodes déchantillonnage. Behavior 49, 227–266. doi: 10.1163 / 156853974×00534
Résumé PubMed | CrossRef Texte intégral | Google Scholar
Anderson, J. R. et Meno, P. (2003). Influences psychologiques sur le bâillement chez les enfants. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J., et Cirulli, E. T. (2014). La variation individuelle de la sensibilité au bâillement contagieux est très stable et largement inexpliquée par lempathie ou dautres facteurs connus. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
Résumé PubMed | CrossRef Texte intégral | Google Scholar
Bretz, F., Hothorn, T., et Westfall, P. (2010). Comparaisons multiples avec R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Député, BL (1994). Etude éthologique du bâillement chez les primates. I. Analyse quantitative et étude de la causalité chez deux espèces de singes de lAncien Monde (Cercocebus albigena et Macaca fascicularis). Ethology 98, 221–245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Texte intégral | Google Scholar
Dobson, A. J. (2002). Une introduction aux modèles linéaires généralisés, 2e éd. Boca Raton, FL: Chapman et Hall.
Google Scholar
Katsuki, F., et Constantinidis, C (2014). Attention ascendante et descendante: processus différents et systèmes neuronaux qui se chevauchent. Neuroscientist 20, 509–521. doi: 10.1177 / 1073858413514136
Résumé PubMed | CrossRef Texte intégral | Google Scholar
Singer, T. (2006). La base neuronale et lontogénie de lempathie et de la lecture mentale: revue de la littérature et implications pour les recherches futures. Neurosci. Biobehav. Rev. 30, 855–863. doi: 10.1016 / j.neubiorev.2006.06.011
Résumé PubMed | CrossRef Texte intégral | Google Scholar
Statistique Canada (2009). Catégories dâge, regroupements de cycles de vie. Ottawa: Statistique Canada.
Google Scholar